

Tre caratteristiche e funzioni nell'organismo
- 1042
- 165
- Benedetta Rinaldi
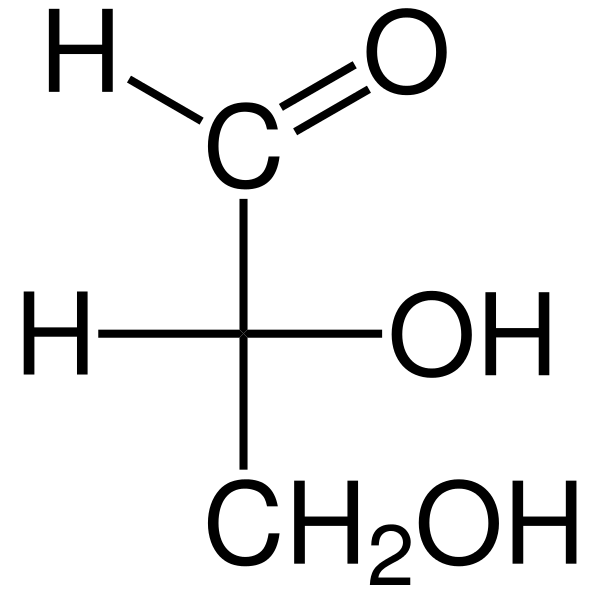
IL Triose Sono monosaccaridi a tre carbonio la cui formula chimica empirica è C3H6O6. Ci sono due triose: gliceraldeide. Le triose sono importanti nel metabolismo perché collegano tre vie metaboliche: glicolisi, gluconeogenesi e percorso del fosfato pentosio.
Durante la fotosintesi, il ciclo di Calvin è una fonte di triose che servono per la biosintesi di fruttosio-6-fosfato. Questo zucchero, in modo fosforilato, viene convertito da passaggi con catalizzazione enzimaticamente in polisaccaridi di riserva o strutturali.
Fonte: Wesalius [dominio pubblico] Le triose partecipano alla biosintesi dei lipidi che fanno parte delle membrane cellulari e degli adipociti.
[TOC]
Caratteristiche
La gliceraldeide aldosa ha un atomo di carbonio quirale e, quindi, ha due enantiomeros, la L-gliceraldeide e la d-gliceraldeide. Entrambi gli enantiomeri D e L hanno caratteristiche chimiche e fisiche diverse.
La d-gliceraldeide ha rotto il piano della luce polarizzata a destra (+) e ha una rotazione [α] D, a 25 ° C, da +8.7 °, mentre la L-glicecentelide ha rotto il piano della luce polarizzata a sinistra (-) e ha una rotazione [α] D, a 25 ° C, da -8.7 °.
Il carbonio chirale di gliceraldeide è carbonio 2 (C-2), che è un alcol secondario. La proiezione di Fischer rappresenta il gruppo idrossilico (-OH) della gliceraldeide D a destra e il gruppo OH della L-Glyce sinistra.
Diidrossiacetone manca di carboni chirali e non ha forme enantiomeriche. L'aggiunta di un gruppo di idrossimetilene (-Choh) a gliceraldeide o diidrossicetone consente la creazione di un nuovo centro chirale. Di conseguenza, lo zucchero è una tetrosa perché ha quattro carboni.
L'aggiunta di un gruppo -chah a Terosa crea un nuovo centro chirale. Lo zucchero formato è un pentosio. Gruppi -Chah può continuare ad essere aggiunti a un massimo di dieci carboni.
Può servirti: filogenesiFunzioni nell'organismo
Le triose come intermediari nella glicolisi, nella gluconeogenesi e nel percorso del pentosa fosfato
La glicolisi consiste nella rottura della molecola di glucosio in due molecole di piruvato per produrre energia. Questa via implica due fasi: 1) fase preparatoria o consumo di energia; 2) Fase di generazione di energia. Il primo è quello che produce le triose.
Nella prima fase, il contenuto energetico privo di glucosio viene aumentato, dalla formazione di fosfoéster. In questa fase, il trifosfato di adenosina (ATP) è il donatore di fosfato. Questa fase culmina con la conversione del fosfoé di fruttosio 1.6-bifosfato (F1.6bp) in due triosas fosfato, gliceraldeide 3-fosfato (GA3P) e diidroxiacetone-fosfato (DHAP).
La gluconeogenesi è la biosintesi del glucosio dal piruvato e altri intermediari. Usa tutti gli enzimi di glicolisi che catalizzano le reazioni la cui variazione di energia standard biochimica è in equilibrio (ΔGº ~ 0). Per questo motivo, la glicolisi e la gluconeogenesi hanno intermediari comuni, tra cui GA3P e DHAP.
Il percorso del pentosio fosfato è costituito da due stadi: una fase ossidativa di glucosio-6-fosfato e un'altra formazione di NADPH e ribosa-5-fosfato. Nella seconda fase, il ribosio a 5-fosfato viene convertito in intermediari di glicolisi, F1.6BP e GA3P.
Le triose e il ciclo di Calvin
La fotosintesi è divisa in due fasi. Nel primo si verificano reazioni dipendenti dalla luce che producono NADPH e ATP. Queste sostanze sono usate nel secondo, in cui vi è fissazione di anidride carbonica e formazione esososa dalle triose attraverso un percorso noto come ciclo di Calvin.
Nel ciclo di Calvin, l'enzima ribulosio 1,5-bifario-ossigenasi (Rubisco) catalizza l'unione covalente del CO2 Al pentosio ribulosa 1.5-bifosfato e rompe l'intermediario instabile di sei atomi di carbonio in due molecole di tre atomi di carbonio: il 3-fosfoglicerato.
Può servirti: oligosaccaridi: caratteristiche, composizione, funzioni, tipiAttraverso reazioni enzimatiche che includono la fosforilazione e la riduzione del 3-fosfoglicerato, usando ATP e NADP, si verifica GA3P. Questo metabolita viene convertito in fruttosio 1.6-bifosfato (F1.6bp) da una via metabolica simile alla gluconeogenesi.
Per azione di una fosfatasi, F1.6bp viene convertito in fruttosio-6-fosfato. Quindi, un fosfhexose isomease produce glucosio 6-fosfato (GLC6P). Infine, un'epicherasi converte GLC6P in 1-fosfato di glucosio, che serve per la biosintesi dell'amido.
Le triose e i lipidi delle membrane biologiche e adipocita
Ga3p e Dhap possono formare glicerolo fosfato che è un metabolita necessario per la biosintesi di triacilgliceroli e glicerolipidi. Questo perché entrambe le Triosas fosfato possono essere interconvertite da una reazione catalizzata dalla triosa isomerasi fosfato, che mantiene entrambe le triose in equilibrio.
L'enzima deidrogenasi glicerolo-fosfato catalizza una reazione di riduzione ossida, in cui il NADH dona un paio di elettroni al DHAP per formare glicerolo 3-fosfato e NAD+. Il 3-fosfato L-glicerolo fa parte dello scheletro dei fosfolipidi che sono parte strutturale delle membrane biologiche.
Il glicerolo è proquirale, manca di carboni asimmetrici, ma quando uno dei suoi due alcoli primari forma un fosfoester, può essere correttamente chiamato 3-fosfato L-glicerolo o 3-fosfato.
I glyceofosfolipidi sono anche chiamati fosfogliceridi, nominati come acido fosfatidico derivato. I fosfogliceridi possono formare fosfoacilgliceroli formando legami estere con due acidi grassi. In questo caso, il prodotto risultante è 1,2-fosfodiacilglicerolo, che è un componente importante delle membrane.
Una gliceofosfasi catalizza l'idrolisi del gruppo di glicerolo fosfato 3-fosfato, producendo glicerolo più fosfato. Il glicerolo può fungere da metabolita iniziale per la biosintesi dei triacilgliceridi, che sono comuni negli adipociti.
Può servirti: test di coagulasi: fondazione, procedura e usiLe triose e le membrane degli archeobatteri
Simile a eubatteri ed eucarioti, il glicerolo 3-fosfato è formato da Triosas fosfato (Ga3p e DHAP). Tuttavia, ci sono differenze: il primo è che il glicerolo 3-fosfato nelle membrane di archeobatteri è L-configurazione, mentre nelle membrane eubatteri e eucariotiche è configurazione D.
Una seconda differenza è che le membrane di archeobatteri formano collegamenti estere con due lunghe catene di idrocarburi di gruppi isoprenoidi, mentre in eubacteria ed eucarioti il glicerolo forma legami estere (1.2-diacilglicerolo) con due catene idrocarbonali di acidi grassi.
Una terza differenza è che, nelle membrane di archeobatteri, le sostituzioni nel fosfato e nel glicerolo 3-fosfato sono diversi da quelli di eubatteri ed eucarioti. Ad esempio, il gruppo fosfato è collegato al disaccaride α-Glucopiranso- (1®2)-β-Galatofuranosa.
Riferimenti
- Cui, s. W. 2005. Carboidrati alimentari: chimica, proprietà fisiche e applicazioni. CRC Press, Boca Raton.
- Cock, p., Mäkinen, K, Honkala e., Saag, m., Kennepohl, e., Eapes, a. 2016. L'eritritolo è più efficace dello xilitolo e del sorbitolo nella gestione degli endpoint di salute orale. International Journal of Dentistry.
- Nelson, d. L., Cox, m. M. 2017. Principi di biochimica Lechinger. W. H. Freeman, New York.
- Sinnott, m. L. 2007. Struttura e meccanismo di chimica e biochimica dei carboidrati. Royal Society of Chemistry, Cambridge.
- Stick, r. V., Williams, s. J. 2009. Carboidrati: le molecole essenziali della vita. Elsevier, Amsterdam.
- Voet, d., Voet, j. G., Pratt, c. W. 2008. Fondamenti di biochimica - vita a livello molecolare. Wiley, Hoboken.
- « Caratteristiche dell'aldosio, tipi, numero di carboni
- Fluoxetina (prozac) meccanismo d'azione ed effetti collaterali »