

Caratteristiche, funzioni, componenti dei pori nucleari
- 1150
- 105
- Rosolino Santoro
Lui poro nucleare (Dal greco, pori = passo o transito) è la "porta" del nucleo che consente la maggior parte del trasporto tra nucleoplasma e citoplasma. Il poro nucleare si unisce alle membrane interne ed esterne del nucleo per formare canali, che servono per il trasporto di proteine e RNA.
La parola Poro non riflette la complessità della struttura in questione. Per questo motivo, è preferibile fare riferimento al complesso dei pori nucleari (CPN), anziché al poro nucleare. Il CPN può sperimentare cambiamenti nella sua struttura durante il trasporto o gli stati del ciclo cellulare.
Recentemente, è stato scoperto che le nucleoporine, le proteine che compongono il CPN, hanno un ruolo importante nella regolazione dell'espressione genica. Pertanto, quando si verificano mutazioni che influenzano la funzione delle nucleoporine, le patologie sono prodotte nell'uomo, come autoimmune, cardiomiopatie, infezioni virali e cancro.
[TOC]
Caratteristiche
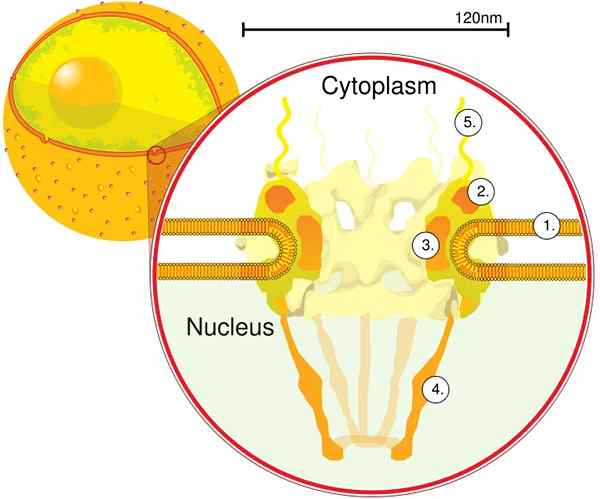
Attraverso la tomografia elettronica è stato determinato che il CPN ha uno spessore di 50 nm, un diametro esterno tra 80 e 120 nm e un diametro interno di 40 nm. Grandi molecole come la grande subunità di ribosomi (PM 1590 kDa) possono essere esportate al di fuori del nucleo attraverso il CPN. Si stima che ci siano tra 2000 e 4000 cpn per nucleo.
Il peso molecolare di un singolo CPN è approssimativamente tra 120 e 125 MDA (1 MDA = 106 Da) nei vertebrati. Al contrario, il CPN è più piccolo nei lieviti, in cui ha circa 60 MDA. Nonostante le enormi dimensioni del CPN, le nucleoporine sono altamente conservate in tutti gli eucarioti.
La traslocalizzazione attraverso il CPN è un processo rapido, la cui velocità è 1000 traslocazioni/secondo. Tuttavia, il CPN non determina la direzione del flusso di trasporto stesso.
Questo dipende dal gradiente Rangtp, che è maggiore nel nucleo che nel citosol. Questo gradiente è mantenuto da uno scambiatore di guanine Ran.
Durante il ciclo cellulare, il CPNS sperimenta l'assemblaggio e i cicli di disunità. L'assemblaggio avviene all'interfaccia e immediatamente dopo la mitosi.
Funzioni
L'acido ribonucleico (piccolo RNA nucleare, RNA di messaggero, RNA di trasferimento), proteine e ribonucleoproteine (RNP) deve essere attivamente trasportato tramite CPN. Ciò significa che è richiesta l'energia di ATP e idrolisi GTP. Ogni molecola viene trasportata in modo specifico.
In generale, le molecole di RNA sono piene di proteine che formano complessi RNP, esportati in questo modo. Al contrario, le proteine che vengono attivamente trasportate nel nucleo devono avere un segno di posizione di sequenza nel nucleo (SLN), possessore dei rifiuti di aminoacidi con carico positivo (ad esempio, KKKRK).
Può servirti: mesosomaLe proteine che vengono esportate nel nucleo devono avere un segnale di esportazione al nucleo (NES) ricco di aminoacido leucina.
Oltre a facilitare il trasporto tra il nucleo e il citoplasma, i CPN sono coinvolti nell'organizzazione della cromatina, nella regolazione dell'espressione genica e nella riparazione del DNA. Le nucleoporine (NUPS) promuovono l'attivazione o la repressione della trascrizione, indipendentemente dallo stato di proliferazione cellulare.
Nei lieviti, i nupt si trovano nel CNP dell'involucro nucleare. Nei metazoari sono dentro. Svolgono le stesse funzioni in tutti gli eucarioti.
Importazione di sostanze
Attraverso il CPN c'è una diffusione passiva di piccole molecole in entrambe le direzioni e il trasporto attivo, l'importazione di proteine, l'esportazione di RNA e ribonucleoproteine (RNP) e la navetta delle molecole bidirezionali. Quest'ultimo include RNA, RNP e proteine coinvolte nella segnalazione, nella biogenesi e nella sostituzione.
L'importazione di proteine all'interno del nucleo avviene in due fasi: 1) il legame delle proteine al lato citoplasmatico del CPN; 2) traslocalizzazione dipendente dall'ATP attraverso il CPN. Questo processo richiede l'idrolisi ATP e lo scambio di GTP/PIL tra il nucleo e il citoplasma.
Secondo un modello di trasporto, il complesso proteico ricevente si sposta lungo il canale attraverso l'Unione, la dissociazione e l'Unione alle ripetute sequenze FG delle nucleoporine. In questo modo, il complesso si sposta da una nucleoporina all'altra all'interno del CPN.
Esportazione di sostanze
È simile all'importazione. Ran GTPase impone la direzionalità al trasporto tramite CNP. RAN è un interruttore molecolare con due stati conformazionali, a seconda che sia collegato al PIL o al GTP.
Due proteine regolatorie specifiche innescano la conversione tra i due stati: 1) proteina di attivazione della GTPase citosolica (GAP), che produce idrolisi GTP e quindi converte RAN-GTP in RAN-GDP; e 2) Nuclear Guanina Exchange Factor (GEF), che promuove lo scambio di PIL da parte di GTP e converte RAN-GDP in RAN-GTP.
Il citosol contiene principalmente RAN-GDP. Il nucleo contiene principalmente Ran-GTP. Questo gradiente delle due forme conformazionali di RAN dirige il trasporto nell'indirizzo appropriato.
L'importazione del ricevitore, insieme alla posizione, è facilitata dall'Unione alle ripetizioni-FG. Se raggiungi il lato nucleare del CNP, Ran-GTP si unisce al ricevitore per rilasciare la tua posizione. Pertanto, Ran-GTP crea la direzione del processo di importazione.
L'esportazione nucleare è simile. Tuttavia, Ran-GTP nel nucleo promuove l'unione della posizione al ricevitore di esportazione. Quando il ricevitore di esportazione si muove attraverso il poro verso il citosol, incontra Ran-GAP, che induce l'idrolisi del GTP al PIL. Infine, il ricevitore viene rilasciato dalla sua posizione e RAN-GDP nel citosol.
Può servirti: leucociti polimorfonucleariTrasporto di RNA
L'esportazione di alcuni tipi di RNA è simile all'esportazione di proteine. Ad esempio, ARNT e RNAN (piccolo nucleare) usano il gradiente Rangtp e vengono trasportati attraverso la CPN mediante mezzi. L'esportazione di ribosomi maturi dipende anche dal gradiente Rangtp.
L'mRNA viene esportato in un modo molto diverso dalle proteine e altri RNA. Per la sua esportazione, l'mRNA forma un complesso di messaggero RNP (RNPM), in cui una molecola RNM è circondata da centinaia di molecole proteiche. Queste proteine hanno l'elaborazione, Capping, Empalme e poliadentilly di MNA.
La cellula deve essere in grado di distinguere tra RNPM con RNM maturo e RNPM con mRNA immaturo. L'mRNA, che forma il complesso RPNM, potrebbe adottare topologie che devono essere rimodellate per il trasporto. Prima che l'RNPM entri nel CPN, si verifica una fase di controllo effettuata da complessi proteici di vagabon e esosoma.
Quando viene assemblato RNPM maturo, RPNM viene trasportato attraverso il canale per mezzo di un ricevitore di trasporto (NXF1-NXT1). Questo ricevitore ha bisogno di idrolisi ATP (non un gradiente Rangtp) per stabilire il rimodellamento della direzionalità dell'RNPM, che raggiungerà il citoplasma.
Il complesso dei pori nucleari e il controllo dell'espressione genica
Alcuni studi indicano che i componenti CPN potrebbero influenzare la regolazione dell'espressione genetica attraverso il controllo della struttura della cromatina e la sua accessibilità ai fattori di trascrizione.
Negli eucarioti dell'evoluzione più recente, l'eterocromatina si trova preferibilmente sulla periferia del nucleo. Questo territorio è interrotto dai canali di euchromatina, che sono mantenuti dal cestino nucleare CPN. L'associazione del cestino nucleare con l'euchromatina è correlata alla trascrizione genica.
Ad esempio, l'attivazione della trascrizione a livello di CPN implica l'interazione dei componenti del cestino nucleare con proteine come la saga di acetiltransferasi dell'istone e i fattori di esportazione dell'RNA.
Pertanto, il cestino nucleare è una piattaforma per numerosi geni di mantenimento (Faccende domestiche) altamente trascritto e geni fortemente indotti dai cambiamenti in condizioni ambientali.
Il complesso dei pori nucleari e della virologia
L'infezione da cellule virali di eucariota dipende da CPN. In ogni caso di infezione virale, il suo successo dipende dal DNA, RNA o RPN che attraversa il CPN per raggiungere il suo obiettivo finale, che è la replicazione del virus.
Può servirti: inclusioni citoplasmatiche: cosa sono, caratteristiche, funzioniAPE Virus 40 (SV40) è stato uno dei modelli più studiati per studiare la funzione del CPN nella traslocalizzazione all'interno del nucleo. Questo perché Sv40 ha un piccolo genoma (5.000 basi).
È stato dimostrato che il trasporto del DNA del virus è facilitato le proteine proteiche del virus, che proteggono il virus fino a raggiungere il nucleo.
Componenti
Il CPN è incorporato all'interno dell'involucro nucleare ed è costituito da circa 500 e 1000 nupt. Queste proteine sono organizzate in sottocompleci o moduli strutturali, che interagiscono tra loro.
Il primo modulo è un componente centrale, o anello, all'interno del poro a forma di un orologio di sabbia, che è limitato da un altro anello di diametro di 120 nm su entrambi i lati, intranucleare e citoplasmatico. Il secondo modulo sono gli anelli del nucleo e del citoplasma (120 nm di diametro ciascuno) situati attorno al componente a forma di sabbia.
Il terzo modulo sono otto filamenti proiettati dall'anello da 120 nm all'interno del nucleoplasma e forma una struttura a forma di cesto. Il quarto modulo è costituito dai filamenti proiettati al lato del citoplasma.
Il complesso a forma di Y, composto da sei NUP e proteine SEH 1 e sec 13, è il complesso più grande e meglio caratterizzato del CNP. Questo complesso è l'unità essenziale che fa parte delle impalcature CPN.
Nonostante la bassa somiglianza tra le sequenze NUPS, l'impalcatura CPN è altamente conservata in tutti gli eucarioti.
Riferimenti
- Beck, m., Ferito, e. 2016. Il complesso dei pori nucleari: comprendere la sua funzione attraverso l'intuizione strutturale. Recensioni della natura, biologia cellulare molecolare, doi: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Proteine dei pori nucleari e il controllo delle funzioni del genoma. Geni e sviluppo, 29, 337-349.
- Kabachinski, g., Schwartz, t.O. 2015. Il complesso dei pori nucleari - Struttura e funzione a colpo d'occhio. Journal of Cell Science, 128, 423-429.
- Knocknhauer, k.E., Schwartz, t.O. 2016. Il complesso dei pori nucleari come cancello flessibile e dinamico. Cell, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.W. 2014. Il nucleare multifunzionale per complesso: una piattaforma per il controllo dell'espressione genica. Opinione attuale della biologia cellulare, doi: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.P., Steffen, j. 2017. Repubblicazione: il complesso dei pori nucleari: una revisione completa della struttura e della funzione. International Journal of Academic Medicine, 3, 51-59.
- Tran, e.J., WADE, s.R. 2006. Nucleare dinamico per complessi: vita al limite. Cell, 125, 1041-1053.