

Sintesi dei tipi lipidici e dei loro meccanismi principali
- 2708
- 488
- Silvano Montanari
IL Sintesi lipidica È costituito da una serie di reazioni enzimatiche per mezzo delle quali sono condensati idrocarburi a catena corta per formare molecole a catena più lunghe che possono successivamente subire diverse modifiche chimiche.
I lipidi sono un tipo molto vario di biomolecole sintetizzate da tutte le cellule viventi e sono specializzati in molteplici funzioni essenziali per il mantenimento della vita cellulare.
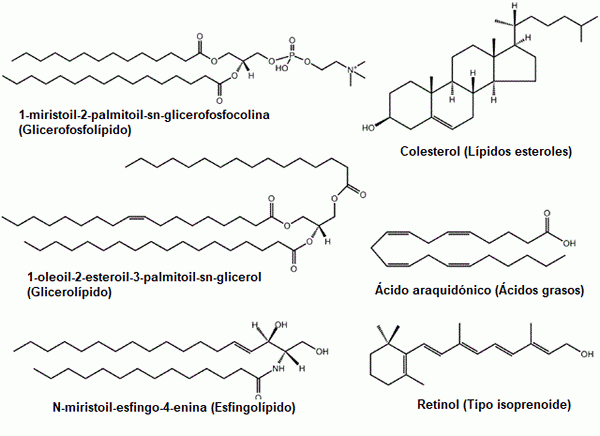 Alcuni esempi di lipidi comuni: glicerofosfolipidi, steroli, glicerolipidi, acidi grassi, sfingolipidi e prenoli (fonte: il caricatore originale era LMAPS in inglese Wikipedia. / GFDL 1.2 (http: // www.gnu.Org/licenze/vecchi licenze/FDL-1.2.html) via Commons, adattato da Raquel Parada)
Alcuni esempi di lipidi comuni: glicerofosfolipidi, steroli, glicerolipidi, acidi grassi, sfingolipidi e prenoli (fonte: il caricatore originale era LMAPS in inglese Wikipedia. / GFDL 1.2 (http: // www.gnu.Org/licenze/vecchi licenze/FDL-1.2.html) via Commons, adattato da Raquel Parada) I lipidi sono i componenti principali delle membrane biologiche, un fatto che le rende molecole fondamentali per l'esistenza di cellule come entità isolate di ciò che li circonda.
Alcuni lipidi hanno anche funzioni specializzate come pigmenti, cofattori, trasportatori, detergenti, ormoni, messaggeri intra-estracellulari, ancore covalenti per proteine di membrana, ecc. Pertanto, la capacità di sintetizzare diversi tipi di lipidi è fondamentale per la sopravvivenza di tutti gli organismi viventi.
Questo folto gruppo di composti è tradizionalmente classificato in diverse categorie o sottogruppi: acidi grassi (saturi e insaturi) lipidi complessi di gliceridi (lipoproteine).
[TOC]
Tipi di lipidi e i loro principali meccanismi di sintesi
Tutte le sequenze di reazione delle rotte di biosintesi lipidica sono endogoniche e riduttivi. In altre parole, usano tutti ATP come fonte di energia e un trasportatore di elettroni ridotto, come NADPH, come potenza riducente.
Successivamente, verranno descritte le principali reazioni delle rotte biosintetiche dei principali tipi di lipidi, ovvero di acidi grassi ed eicosanoidi, triacilgliceroli e fosfolipidi e steroli (colesterolo) (colesterolo).
- Sintesi di acidi grassi
Gli acidi grassi sono molecole estremamente importanti dal punto di vista lipidico, in quanto fanno parte dei lipidi più rilevanti nelle cellule. La sua sintesi, contrariamente a quella che molti scienziati hanno pensato durante i primi studi, non consiste nella via inversa della loro β-ossidazione.
In effetti, questa via metabolica si verifica in diversi compartimenti cellulari e richiede la partecipazione di un intermediario di tre atomi di carbonio noti come malonil-CoA, che non è necessario nell'ossidazione.
 Malonil-Coa. Dominio neuroteker / pub
Malonil-Coa. Dominio neuroteker / pub Inoltre, è strettamente correlato ai gruppi di solfidrile di proteine note come trasportatori di gruppi acilici (ACP, inglese Proteine del vettore acilico).
In generale, la sintesi di acidi grassi, in particolare quello della catena lunga, è un processo sequenziale in cui quattro fasi vengono ripetute in ogni "ritorno" e durante ogni ritorno c'è un gruppo di acido saturo che è il substrato per il prossimo implica un'altra condensa con una nuova molecola malonil-coa.
In ogni ciclo di svolta o di reazione la catena di acido grasso estende due carboni, fino a quando non raggiunge una lunghezza di 16 atomi (palmitato), dopo di che il ciclo lascia.
Formazione malonil-CoA
Questo intermediario di tre atomi di carbonio si forma irreversibilmente da acetil-CoA grazie all'azione di un enzima acetil-COA carbossilasi, che ha un gruppo prostatico di biotina che è covalentemente collegato all'enzima e che partecipa a questa catalisi in questa catalisi in due passi in due passi.
In questa reazione, un gruppo carbossilico derivato da una molecola di bicarbonato (HCO3-) viene trasferito nella biotina in una forma dipendente dall'ATP, in cui il gruppo biotinile svolge la funzione del "trasportatore temporaneo" della molecola mentre trasferisce a acetil-COA, producendo malonil-coa.
Può servirti: test ossidasi: fondazione, procedura e usiNella sequenza di sintesi di acidi grassi, l'agente riducente utilizzato è il NADPH e i gruppi attivanti sono due gruppi TIOL (-sh) che fanno parte di un complesso multienzimatico chiamato acido grasso sintasi, che è quello più importante nella catalisi sintetica sintetica.
Nei vertebrati, il complesso di acidi grassi sintasi fa parte di una singola grande catena polipeptidica, in cui sono rappresentate le 7 attività enzimatiche caratteristiche della via della sintesi, nonché l'attività idrolitica necessaria per rilasciare intermediari alla fine della sintesi.
 Struttura enzimatica degli acidi grassi Sintasa (fonte: Boehringer Ingelheim/CC BY-SA (https: // creativeCommons.Org/licenze/by-sa/4.0) via Wikimedia Commons)
Struttura enzimatica degli acidi grassi Sintasa (fonte: Boehringer Ingelheim/CC BY-SA (https: // creativeCommons.Org/licenze/by-sa/4.0) via Wikimedia Commons) Le 7 attività enzimatiche di questo complesso sono: proteina di trasporto del gruppo Acilo (ACP), acetil-CoA-ACP Transacetilas cetoacil-ACP reduttasi (KR), β-idrossicile-ACP dehydratasa (HD) e rabbia-ACP reductasi (ER).
Prima che le reazioni di condensazione possano verificarsi per assemblare la catena di acido grasso, i due gruppi di Tiol nel complesso enzimatico vengono "caricati" con i gruppi acilici: prima di tutto, un acetil -coa viene trasferito al gruppo -sh di una cisteina Nella sintasi β-cethoacil-ACP del complesso, reazioni catalizzate dall'enzima transacetila acetil-CoA-ACP (AT) (AT).
Successivamente, un gruppo malonico viene trasferito da una molecola malonil -CoA al gruppo -sh del trasporto ACILO Group (ACP) fa parte del complesso di acidi grassi sintasi.
La sequenza di quattro reazioni per ogni "ritorno" del ciclo di reazione è la seguente:
- Condensa: i gruppi acetilici e malonilici "caricati" nell'enzima sono condensati per formare una molecola acetoacetil -acp, che è collegata alla porzione ACP attraverso un gruppo -sh. In questo passaggio c'è una molecola di CO2 ed è catalizzata dal β-zoacil-ACP sintasi (il gruppo acetilico occupa la posizione di "metil terminale" del complesso acetoacetil-ACP).
- Riduzione del gruppo carbonilico: il gruppo carbonile nella posizione C3 dell'acetoacetil-ACP è ridotto per formare D-β-idrossibutilil-ACP, catalizzata dalla reazione dalla β-cethoacil-ACP reduttasi, che NADPH utilizza come donatore elettronico.
- Disidratazione: i carboni C2 e C3 del d-β-idrossibutilil-ACP sono privi di molecole d'acqua, formando un doppio legame che termina con la produzione del nuovo composto trans-∆2-butenoil-acp. Questo processo è mediato da un enzima β-idrossil-ACP deidratasi (HD) (HD).
- Riduzione del doppio legame: il doppio legame del composto formato nella fase di disidratazione è saturo (ridotto) per dare origine a butiril-ACP dalla reazione catalizzata dall'enzima rabbia-ACP reduttasi (ER), che utilizza anche NADPH come agente riducente.
The synthesis reactions occur until a palmitate molecule (16 carbon atoms) is formed, which is hydrolyzed of the enzymatic complex and released as a possible precursor for fatty acids of greater length chains, which are produced by elongation systems of fatty acids located in the smooth porzione del reticolo endoplasmatico e nei mitocondri.
Può servirti: fauna e flora del mare peruvianoLe altre modifiche che queste molecole possono soffrire come le deaturazioni, ad esempio, sono catalizzate da diversi enzimi, che generalmente si verificano nel reticolo endoplasmatico liscio.
- Sintesi di eicosanoidi
Gli eicosanoidi sono lipidi cellulari che hanno funzioni come molecole di messaggero "corta", prodotte da alcuni tessuti per comunicare con i loro tessuti vicini. Queste molecole sono sintetizzate da acidi grassi polinsaturi di 20 atomi di carbonio.
Prostaglandine
In risposta a uno stimolo ormonale, l'enzima fosfolipasi attacca i fosfolipidi di membrana e rilascia Araquidonato dal carbonio glicerolo 2. Questo composto viene convertito in prostaglandine grazie a un enzima del reticolo endoplasmatico liscio con attività bifunzionale: cicloossigenasi (COX) o prostaglandina H2 sintasi.
Tromboxan
Le prostaglandine possono essere convertite in trombossani grazie alla sintasi trombossana nelle piastrine del sangue (trombociti). Queste molecole partecipano alle fasi iniziali della coagulazione del sangue.
- Sintesi di triacilgliceroli
Gli acidi grassi sono molecole fondamentali per la sintesi di altri composti più complessi nelle cellule, come i triacilgliceroli o la membrana lipidi glicerofosfolipidi lipidici (processi che dipendono dalle esigenze metaboliche cellulari).
Gli animali producono triacilgliceroli e glyceofosfalipidi da due precursori comuni: l'acile grasso-COA e il l-glicerolo 3-fosfato. L'acilico di grasso-CoA è prodotto da sintesi acil-CoA che partecipano all'ossidazione β, mentre L-glicerolo 3-fosfato è ottenuto dalla glicolisi e dall'azione di due enzimi alternativi: glicerolo deidrogenasi 3-fosfato.
I triacilgliceroli sono formati dalla reazione tra due molecole di FLO-CoA Acil e una molecola di diacilglicerolo 3-fosfato; Queste reazioni di trasferimento sono catalizzate da transferasi specifici.
In this reaction there is, initially, phosphatidic acid, which is dephosphorylated by an enzyme phosphatide acid phosphatase to produce 1.2-Diacylglycerol, which is capable, again, of accepting a third molecule of fat-coa acyl, producing triacilglyceroll.
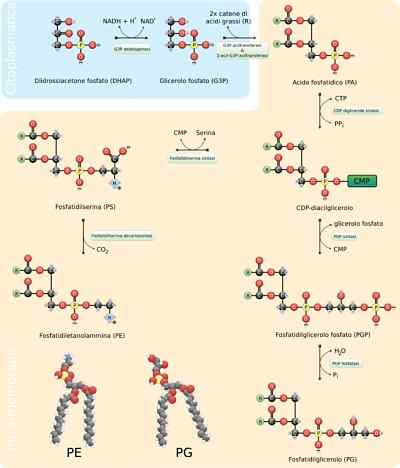
- Sintesi di fosfolipidi
I fosfolipidi sono molecole estremamente variabili, poiché molti diversi possono formarsi mediante la combinazione di acidi grassi e gruppi "testa" con lo scheletro di glicerolo (glyceofosfolipidi) o sfinosina (sfingolipidi) che li caratterizzano.
L'assemblaggio generale di queste molecole richiede la sintesi dello scheletro di glicerolo o sferico, dell'unione con gli acidi grassi corrispondenti, sia per sterilizzazione che per amidazione, l'aggiunta di un gruppo "testa" idrofilo attraverso un legame con fosfoditester e, se necessario , L'alterazione o lo scambio di questi ultimi gruppi.
Negli eucarioti questo processo si verifica nel reticolo endoplasmatico liscio e anche nella membrana mitocondriale interna, dove possono rimanere indefinitamente o da dove possono essere traslocati in altri luoghi.
Passi di reazione
I primi passi della reazione di sintesi dei glicerofosfolipidi sono equivalenti a quelli della produzione di triacilgliceroli, poiché una molecola di glicerolo a 3 fosfato viene esterificata in due molecole di acidi grassi nei carboni 1 e 2, formando acido fosfatidico. È comune trovare fosfolipidi che hanno acidi grassi saturi nel C1 e insaturi nel C2 del glicerolo.
Può servirti: resistenzaL'acido fosfatidico può anche essere prodotto mediante fosforilazione di una molecola di diacilglicerolo già sintetizzata o "riciclata".
I gruppi polari "head" di queste molecole sono formati attraverso collegamenti di fosfodié. La prima cosa che dovrebbe accadere affinché questo processo sia correttamente è la "attivazione" di uno dei gruppi idrossilici che partecipa al processo per mezzo di idrossile che partecipa alla reazione.
Se questa molecola si lega al diacilglicerolo, si forma allora CDP-diacilglicerolo (la forma "attivata" dell'acido fosfatidico), ma ciò può accadere anche sul gruppo idrossile della "testa" del gruppo ".
Nel caso della fosfatidilserina, ad esempio, la diacilglicerolo è attivata dalla condensazione della molecola di acido fosfatidico con una molecola di citidina TRIF-citidina (CTP), che forma CDP-diacilglicerolo ed eliminando un pirofosfato.
Se una molecola MMP (citidina monofosfato) si muove attraverso un attacco nucleofilo della serina o dell'idrossile idrossile nel carbonio 1 del glicerolo 3-fosfato. Fosfato produce fosfatidilglicerolololololololololololololololololo.
Entrambe le molecole prodotte in questo modo servono come precursori per altri lipidi di membrana, che spesso condividono le vie biosintetiche l'una dall'altra.
- Sintesi del colesterolo
Il colesterolo è una molecola essenziale per gli animali che possono essere sintetizzati dalle sue cellule, quindi non è essenziale nella dieta quotidiana. Questa molecola di 27 atomi di carbonio è prodotta da un precursore: acetato.
Questa molecola complessa è formata da acetil-CoA in quattro fasi principali:
- Condensa di tre unità di acetato per formare il Mevalonato, una molecola intermedia di 6 carboni (prima è formata una molecola di acetoacetil-CoA con due acetil-CoA (enzima di tiolasi) e poi un'altra di β-idrossi-β-metalglutario-COA (HMG- COA) (enzima sinteta HMG-CoA). Il Mevalonato è formato dall'HMG-CoA e grazie all'enzima HMG-CoA reductasi.
- Conversione di Mevalonato in unità isoprene. I primi 3 gruppi di fosfato vengono trasferiti da 3 molecole ATP a Mevalonato. Uno dei fosfati è perso insieme al gruppo carbonile adiacente ed è formato
- Polimerizzazione o condensa di 6 unità isoprene di 5 atomi di carbonio per formare gli atomi a 30 carbonio (una molecola lineare) rasatura (una molecola lineare).
- Cicration of the Excualean per formare i 4 anelli del nucleo steroideo del colesterolo e successivi cambiamenti chimici: ossidazioni, migrazione e eliminazione di gruppi metilici, ecc., Ciò che produce colesterolo.
Riferimenti
- Garrett, r. H., & Grisham, C. M. (2001). Principi di biochimica: con un focus umano. Brooks/Cole Publishing Company.
- Murray, r. K., Granner, d. K., Mayes, p. A., & Rodwell, V. W. (2014). Biochimica illustrata di Harper. McGraw-Hill.
- Nelson, d. L., Lechinger, a. L., & Cox, m. M. (2008). Principi di biochimica Lechinger. Macmillan.
- Jacquemyn, J., Cascaho, a., & Goodchild, R. E. (2017). I dettagli del reticolo endoplasmatico - biosintesi lipidica controllata. EMBO Reports, 18 (11), 1905-1921.
- Ohlrogge, J., & Sfoglia, J. (1995). Biosintesi lipidica. La cella vegetale, 7 (7), 957.
- « José López Portillo Biografia, governo e contributi
- Giungla tropicale nelle caratteristiche del Messico, posizione, sollievo, flora, fauna »