

Stadi, funzioni e inibitori ossidativi
- 783
- 106
- Dott. Rodolfo Gatti
IL fosforilazione ossidativa È un processo in cui le molecole ATP sono sintetizzate da ADP e PYo (Fosfato inorganico). Questo meccanismo è eseguito da batteri e cellule eucariotiche. Nelle cellule eucariotiche, la fosforilazione viene eseguita nella matrice mitocondriale di cellule non fotosintetiche.
La produzione di ATP è diretta dal trasferimento di elettroni dai coenzimi NADH o FADH2 pure2. Questo processo rappresenta la massima produzione di energia nella cellula ed è derivato dalla degradazione di carboidrati e grassi.
Fonte: robot8a [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)] L'energia immagazzinata nei gradienti di carico e pH, noto anche come forza motoria protonica, consente di eseguire questo processo. Il gradiente di proton che viene generato, fa sì che la parte esterna della membrana abbia un carico positivo a causa della concentrazione di protoni (H+) e la matrice mitocondriale è negativa.
[TOC]
Dove si verifica la fosforilazione ossidativa?
Il trasporto di elettroni e i processi di fosforilazione ossidativa sono associati a una membrana. In procarioti questi meccanismi vengono eseguiti attraverso la membrana plasmatica. Nelle cellule eucariotiche sono associati alla membrana dei mitocondri.
Il numero di mitocondri trovati nelle cellule varia in base al tipo di cellula. Ad esempio, nei mammiferi gli eritrociti mancano di questi organelli, mentre altri tipi di cellule, come le cellule muscolari, possono averne milioni.
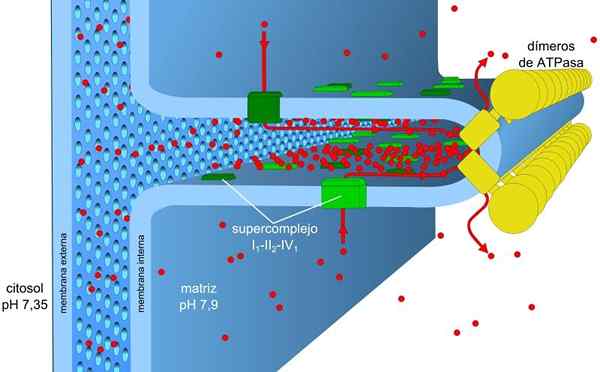
La membrana mitocondriale è costituita da una semplice membrana esterna, una membrana interna un po 'più complessa, e nel mezzo di esse lo spazio intermembrano, dove si trovano molti enzimi che dipendono dall'ATP.
La membrana esterna contiene una proteina chiamata porina che forma i canali per la semplice diffusione di piccole molecole. Questa membrana è responsabile del mantenimento della struttura e della forma dei mitocondri.
La membrana interna ha una maggiore densità ed è ricca di proteine. È anche impermeabile per le molecole e gli ioni, quindi, attraversarlo, hanno bisogno di proteine intermembraniche che le trasportano.
All'interno della matrice, si estendono le pieghe della membrana interna, formando creste che gli consentono di avere una vasta area in un piccolo volume.
Energia cellulare Central
I mitocondri sono considerati come il centro energetico cellulare. In esso ci sono gli enzimi coinvolti nei processi del ciclo dell'acido citrico, l'ossidazione degli acidi grassi e gli enzimi redox e le proteine del trasporto di elettroni e della fosforilazione del dottorato.
Il gradiente di concentrazione dei protoni (gradiente di pH) e il gradiente di carichi o potenziale elettrico nella membrana interna dei mitocondri, sono la causa della forza motoria prottonica. La piccola permeabilità della membrana interna per gli ioni (oltre a H+) consente ai mitocondri di avere un gradiente di tensione stabile.
Trasporto elettronico, pompaggio dei protoni e ATP che ottengono contemporaneamente nei mitocondri, grazie alla forza motoria protonica. Il gradiente di pH mantiene le condizioni acide nella matrice intermembrana e mitocondriale con condizioni alcaline.
Per ogni due elettroni trasferiti a O2 Circa 10 protoni vengono pompati dalla membrana, creando un gradiente elettrochimico. L'energia staccata in questo processo viene prodotta gradualmente passando elettroni attraverso la catena del trasporto.
Fasi
L'energia rilasciata durante le reazioni di riduzione dell'ossido di NADH e FADH2 È considerevolmente alto (circa 53 kcal/mol per ogni coppia di elettroni), quindi da utilizzare nella produzione di molecole ATP, deve essere gradualmente prodotto con il passaggio degli elettroni attraverso i trasportatori.
Questi sono organizzati in quattro complessi situati nella membrana mitocondriale interna. L'accoppiamento di queste reazioni alla sintesi di ATP viene effettuato in un quinto complesso.
Può servirti: linfociti B: caratteristiche, struttura, funzioni, tipiCatena di trasporto di elettroni
NADH trasferisce un paio di elettroni che entrano nel complesso I della catena di trasporto degli elettroni. Gli elettroni vengono trasferiti nel mononucleotide di Flavina e quindi all'ubiquinone (coenzima Q) attraverso un trasportatore di zucchero di ferro. Questo processo rilascia una grande quantità di energia (16,6 kcal/mol).
Ubiquinona trasporta elettroni attraverso la membrana al complesso III. In questo complesso gli elettroni passano attraverso i citocromi B e C1 Grazie a un trasportatore di zuccheri di ferro.
Dal complesso III, gli elettroni passano al complesso IV (citocromo C ossidasi), trasferiti uno dal citocromo C (proteina della membrana periferica). Nel complesso IV gli elettroni passano attraverso una coppia di ioni di rame (CuA2+), quindi al citocromo cA, poi a un'altra coppia di ioni di rame (CuB2+) e da questo al citocromo a3.
Infine, gli elettroni vengono trasferiti a O2 che è l'ultimo accettore e forma una molecola d'acqua (h2O) per ogni coppia di elettroni ricevuti. Il passaggio degli elettroni dal complesso IV a O2 Genera anche molta energia libera (25,8 kcal/mol).
Succinato Coq reduttasi
Il complesso II (coq reduttasi succinato) riceve una coppia di elettroni dal ciclo dell'acido citrico, a causa dell'ossidazione di una molecola di succinato a fumarato. Questi elettroni vengono trasferiti sulla moda, quindi passano attraverso un gruppo di ferro-azufre, all'Ubiquinona. Da questo coenzima vanno al complesso III e seguono il percorso precedentemente descritto.
L'energia rilasciata nella reazione di trasferimento di elettroni alla moda non è sufficiente per aumentare i protoni attraverso la membrana, in modo che in questa fase della catena non vi sia alcuna forza motoria protttonica, e di conseguenza il FADH produce meno H+ che il NADH.
Accoppiamento energetico o trasduzione
L'energia generata nel processo di trasporto degli elettroni precedentemente descritto, dovrebbe essere utilizzata per la produzione di ATP, le reazioni catalizzate dall'ATP sintasi o dall'enzima complesso V. La conservazione di tale energia è nota come accoppiamento energetico e il meccanismo è stato difficile da caratterizzare.
Sono state descritte diverse ipotesi per descrivere questa trasduzione di energia. Il meglio accettato è l'ipotesi di accoppiamento chemosmotico, descritto di seguito.
Accoppiamento chemiosmotico
Questo meccanismo propone che l'energia utilizzata per la sintesi di ATP proviene da un gradiente protonico nelle membrane cellulari. Questo processo interviene in mitocondri, cloroplasti e batteri ed è collegato al trasporto di elettroni.
I complessi I e IV del trasporto elettronico agiscono come pompe protoniche. Questi soffrono di cambiamenti conformazionali che consentono loro di pompare i protoni nello spazio intermembrano. Nel complesso IV per ogni coppia di elettroni, due protoni vengono pompati fuori dalla membrana e altri due rimangono nella matrice che formano H2O.
Ubiquinona nel complesso III accetta protoni di complessi I e II e li libera fuori dalla membrana. I complessi I e III consentono ciascuno il passaggio di quattro protoni per ogni coppia di elettroni trasportati.
La matrice mitocondriale ha una bassa concentrazione di protoni e potenziale elettrico negativo mentre lo spazio intermembrano presenta le condizioni inverse. Il flusso di protoni attraverso questa membrana implica il gradiente elettrochimico che memorizza l'energia necessaria (± 5 kcal/mol per protone) per la sintesi di ATP.
Sintesi ATP
L'enzima della sintetasi ATP è il quinto complesso coinvolto nella fosforilazione ossidativa. È responsabile di trarre vantaggio dall'energia del gradiente elettrochimico per formare ATP.
Questa proteina transmembraale è costituita da due componenti: F0 e f1. Il componente f0 consente il ritorno dei protoni alla matrice mitocondriale che funziona come canale e F1 catalizza la sintesi dell'ATP tramite ADP e PYo, usando l'energia di detto ritorno.
Può servirti: proteine del trasportatore di membrana: funzioni e tipiIl processo di sintesi ATP richiede un cambiamento strutturale in F1 e l'assemblaggio dei componenti f0 e f1. La traslocazione di protoni attraverso F0 Provoca cambiamenti conformazionali in tre subunità di F1, permettendo questo di fungere da motore di rotazione, dirigendo la formazione dell'ATP.
La subunità responsabile dell'Unione di ADP con PYo passa da uno stato debole (L) a un attivo (t). Quando si forma l'ATP, una seconda subunità passa a uno stato aperto (O) che consente il rilascio di questa molecola. Dopo che l'ATP è stato rilasciato, questa subunità passa dallo stato aperto a uno stato inattivo (L).
Le molecole ADP e PYo Si uniscono a una subunità che è passata da uno stato o dallo stato L.
Prodotti
La catena del trasportatore di elettroni e la fosforilazione producono molecole ATP. L'ossidazione NADH produce circa 52,12 kcal/mol (218 kJ/mol) di energia libera.
La reazione globale per l'ossidazione di NADH è:
NADH+1⁄2 o2 +H+ ↔ h2O+NAD+
Il trasferimento di elettroni da NADH e FADH2 Si verifica attraverso diversi complessi, consentendo il cambiamento di energia libera ΔG ° di essere diviso in "pacchetti" più piccoli, che sono accoppiati alla sintesi di ATP.
L'ossidazione di una molecola NADH genera la sintesi di tre molecole ATP. Mentre l'ossidazione di una molecola FADH2 È attaccato alla sintesi di due ATP.
Questi coenzimi provengono dai processi di glicolisi e ciclo di acido citrico. Per ogni molecola di glucosio degradata, stanno finendo 36 o 38 molecole ATP, a seconda della posizione delle cellule. Nel cervello e nel muscolo scheletrico 36 ATP sono prodotti mentre 38 ATP sono prodotti nel tessuto muscolare.
Funzioni
Tutti gli organismi, unicellulari e multicellulari, hanno bisogno di un'energia minima nelle loro cellule per svolgere i processi al loro interno e, a sua volta, mantengono funzioni vitali nel corpo completo.
I processi metabolici richiedono l'energia da eseguire. La maggior parte dell'energia utilizzabile è ottenuta dal degrado di carboidrati e grassi. Questa energia deriva dal processo di fosforilazione ossidativa.
Controllo della fosforilazione ossidativa
La velocità di utilizzo dell'ATP nelle cellule, controlla la sintesi della stessa e, a sua volta, a causa dell'accoppiamento della fosforilazione ossidativa con la catena di trasporto elettronico, generalmente regola anche la velocità di trasporto elettronica.
La fosforilazione ossidativa ha un controllo rigoroso che garantisce che l'ATP non sia generato più velocemente di quanto non sia consumato. Ci sono alcuni passaggi nel processo di trasporto degli elettroni e la fosforilazione accoppiata che regolano il tasso di produzione di energia.
Controllo coordinato della produzione di ATP
I principali percorsi di produzione di energia (ATP cellulare) sono la glicolisi, il ciclo di acido citrico e la fosforilazione ossidativa. Il controllo coordinato di questi tre processi regola la sintesi di ATP.
Il controllo della fosforilazione da parte del rapporto di azione di massa ATP dipende dal preciso contributo degli elettroni nella catena di trasporto. Questo a sua volta dipende dalla relazione [NADH]/[NAD+] che è conservato elevato dall'azione della glicolisi e del ciclo dell'acido citrico.
Questo controllo coordinato viene effettuato regolando i punti di controllo della glicolisi (PFK inibiti dal citrato) e il ciclo dell'acido citrico (deidrogenasi piruvato, citrato di cutista, ichitrato deidrogenasi e α-zetoglutarato deidrogenasi).
Controllo da accettatore
Il complesso IV (citocromo c ossidasi) è un enzima regolato da uno dei suoi substrati, ovvero la sua attività è controllata dal citocromo ridotto C (c2+), che a sua volta è in equilibrio con la relazione delle concentrazioni tra [NADH]/[NAD+] e il rapporto di azione di massa di [ATP]/[ADP] + [PYo".
Può servirti: tono: caratteristiche e funzioniPiù alto il [NADH]/[NAD+] e abbassare il [ATP]/[ADP]+[PYo], una maggiore concentrazione sarà citocromo [c2+] e maggiore sarà l'attività del complesso IV. Questo viene interpretato, ad esempio, se confrontiamo gli organismi con diverse attività di riposo e alta attività.
In un individuo con alta attività fisica, il consumo di ATP e quindi la sua idrolisi ad ADP + PYo Sarà molto alto, generando una differenza nel rapporto di azione di massa che provoca un aumento di [C2+e quindi un aumento della sintesi di ATP. In un individuo a riposo si verifica la situazione inversa.
Alla fine, la velocità della fosforilazione ossidativa aumenta con la concentrazione di ADP all'interno dei mitocondri. Questa concentrazione dipende dai traslocatori ADP-ATP responsabili del trasporto di nucleotidi di adenina e PYo Dal citosol alla matrice mitocondriale.
Agenti decacoplant
La fosforilazione ossidativa è influenzata da alcuni agenti chimici, che consentono al trasporto elettronico di continuare senza la fosforilazione dell'ADP, disaccoppiando la produzione e la conservazione dell'energia.
Questi agenti stimolano la velocità di consumo di ossigeno mitocondri in assenza di ADP, causando anche un aumento dell'idrolisi dell'ATP. Agiscono eliminando un intermediario o rompendo uno stato di energia dalla catena del trasportatore di elettroni.
Il 2,4-dinitrofenolo, un acido debole che passa attraverso le membrane mitocondriali, è responsabile della dissipazione del gradiente dei protoni, perché si uniscono a loro sul lato acido e li rilasciano sul lato base.
Questo composto è stato usato come "pillola dimagrante" perché è stato scoperto che ha prodotto un aumento della respirazione, quindi un aumento del tasso metabolico e della perdita di peso associata. Tuttavia, è stato dimostrato che il suo effetto negativo potrebbe persino causare la morte.
La dissipazione del gradiente protonico produce calore. Le cellule del tessuto adiposo marrone usano disaccoppiamento, controllate ormonalmente, per produrre calore. I mammiferi ibernati e i neonati che non hanno i capelli sono costituiti da questo tessuto che li serve come una sorta di coperta termica.
Inibitori
I composti o agenti inibitori impediscono sia il consumo di o2 (trasporto elettronico) come la fosforilazione ossidativa associata. Questi agenti evitano la formazione di ATP attraverso l'uso dell'energia prodotta nel trasporto elettronico. Pertanto, la catena di trasporto si ferma in quanto tale consumo di energia non è disposto.
L'antibiotico oligomicina funziona come un inibitore della fosforilazione in molti batteri, impedendo la stimolazione dell'ADP dalla sintesi di ATP.
Ci sono anche agenti ionofori, che compongono complessi solubili a grasso con cationi come K+ e na+, e passare attraverso la membrana mitocondriale con questi cationi. I mitocondri utilizzano quindi l'energia prodotta nel trasporto elettronico per pompare i cationi invece di sintetizzare ATP.
Riferimenti
- Alberts, b., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, P. (2004). Biologia cellulare essenziale. New York: Garland Science.
- Cooper, g. M., Hausman, r. E. & Wright, n. (2010). La cellula. (pp. 397-402). Marbán.
- Devlin, t. M. (1992). Libro di testo di biochimica: con correlazioni cliniche. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, C. M. (2008). Biochimica. Thomson Brooks/Cole.
- Lodish, h., Darnell, J. E., Berk, a., Kaiser, c. A., Krieger, m., Scott, m. P., & Matsudaira, P. (2008). Biologia cellulare mollecolare. Macmillan.
- Nelson, d. L., & Cox, m. M. (2006). Principi di biochimica Lechinger 4a edizione. Ed Omega. Barcellona.
- Voet, d., & Voet, J. G. (2006). Biochimica. Ed. Pan -American Medical.
- « Caratteristiche di Euglenofita, riproduzione, nutrizione, classificazione
- Formazione, funzione e gruppi di glucosidi »