

Ossidazione beta dei passi, delle reazioni, dei prodotti, della regolazione degli acidi grassi
- 4928
- 585
- Cosetta Messina
IL Ossidazione beta di acidi grassi È la via del catabolismo (degradazione) degli acidi grassi, che ha come funzione principale la produzione o il "rilascio" dell'energia contenuta nei legami di queste molecole.
Questo percorso fu scoperto nel 1904 grazie agli esperimenti condotti dal tedesco Franz Knoop, che consisteva nella somministrazione, ratti sperimentali, di acidi grassi il cui gruppo metilico finale era stato modificato con un gruppo fenilico.
Ossidazione dello schema beta di acidi grassi (fonte: Arturo González Laguna [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)] via Wikimedia Commons) Knoop si aspettava che i prodotti di catabolismo di questi acidi grassi "analogici" seguissero percorsi simili alla via di ossidazione degli acidi grassi normali (i nativi non modificati). Tuttavia, ha scoperto che c'erano differenze nei prodotti ottenuti a seconda del numero di atomi di carbonio degli acidi grassi.
Con questi risultati, Knoop ha proposto che il degrado si è verificato in "passaggi", a partire da un carbonio β "attacco" (posizione 3 rispetto al gruppo carbossilico terminale), rilasciando frammenti di due atomi di carbonio.
Successivamente è stato dimostrato che il processo richiede energia sotto forma di ATP, che si verifica nei mitocondri e che i frammenti di due atomi di carbonio entrano nel ciclo di Krebs come acetil-CoA.
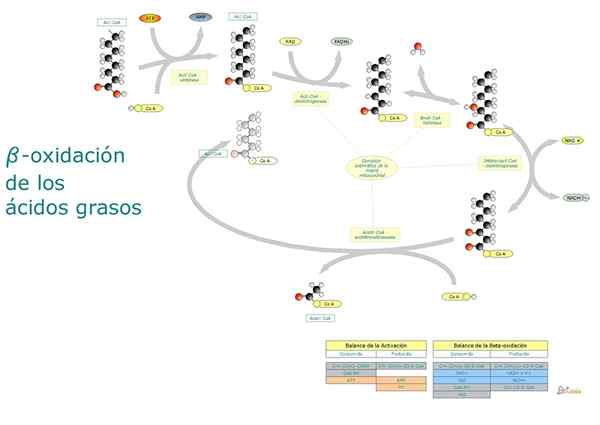
In breve, l'ossidazione beta degli acidi grassi implica l'attivazione del gruppo carbossilico terminale, il trasporto di acidi grassi attivati verso la matrice mitocondriale e l'ossidazione "sfalsata" di due in due carboni dal gruppo carbossilico.
Come molti processi anabolici e catabolici, questa via è regolata, poiché merita la mobilizzazione degli acidi grassi di "riserva" quando le altre rotte cataboliche non sono sufficienti per soddisfare le esigenze di energia cellulare e corpo.
[TOC]
Passi e reazioni
Gli acidi grassi sono prevalentemente in citosol, provengono già da rotte biosintetiche o depositi di grasso che sono immagazzinati da alimenti ingeriti (che devono entrare nelle cellule).
- Attivazione di acidi grassi e trasporto nei mitocondri
L'attivazione di acidi grassi richiede l'uso di una molecola ATP e ha a che fare con la formazione di coniugati di acil tioé con il coenzima a.
Può servirti: importanza della biologiaQuesta attivazione è catalizzata da un gruppo di enzimi chiamati lunghezza specifica di acetil-CoA rispetto alla lunghezza della catena di ciascun grasso. Alcuni di questi enzimi attivano acidi grassi mentre vengono trasportati nella matrice mitocondriale, poiché sono incorporati nella membrana mitocondriale esterna.
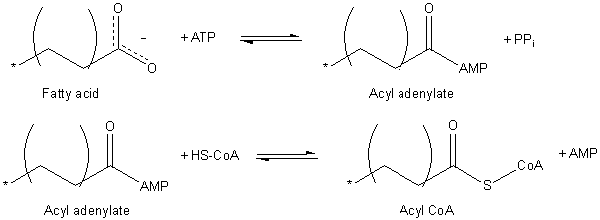 Attivazione di acidi grassi (fonte: JAG123 in inglese Wikipedia [dominio pubblico] via Wikimedia Commons)
Attivazione di acidi grassi (fonte: JAG123 in inglese Wikipedia [dominio pubblico] via Wikimedia Commons) Il processo di attivazione si verifica in due fasi, producendo prima un acile adenilato dall'acido grasso ATP, in cui viene rilasciata una molecola di pirofosfato (PPI). Il gruppo carbossilico attivato dall'ATP viene successivamente attaccato dal gruppo Tiol del coenzima formando Acil-CoA.
La traslocazione Acil-CoA attraverso la membrana mitocondria è raggiunta grazie a un sistema di trasporto noto come navetta di carnitina.
- Ossidazione beta di acidi grassi saturi con una coppia di atomi di carbonio
Il degrado degli acidi grassi è una via ciclica, poiché il rilascio di ciascun frammento di due atomi di carbonio è immediatamente seguito da un altro, fino a raggiungere la lunghezza totale della molecola. Le reazioni che hanno parte in questo processo sono le seguenti:
- Deidrogenazione.
- Idratazione di un doppio legame.
- Deidrogenazione di un gruppo idrossile.
- Frammentazione per l'attacco di una molecola di acetil-CoA sul carbonio β.
Reazione 1: prima deidrogenazione
Consiste nella formazione di un doppio legame tra il carbonio α e il carbonio β mediante eliminazione di due atomi di idrogeno. È catalizzato da un enzima Acil-CoA deidrogenasi, che forma una molecola trans trans.
Reazioni 2 e 3: idratazione e deidrogenazione
L'idratazione è catalizzata dal coot di angeli+.
L'idratazione del bianco trans.
Il FADH2 e il NADH prodotto nelle prime tre reazioni dell'ossidazione beta sono rilESSI.
Può servirti: Ovogonie: cosa sono, descrizione, morfologiaReazione 4: frammentazione
Ogni ciclo beta di ossidazione che elimina una molecola di due atomi di carbonio.
Questa reazione è catalizzata dall'enzima β-cototoolasi o tisolasi e i suoi prodotti sono una molecola Acil-CoA (l'acido grasso attivato con due atomi di carbonio in meno) e uno di acetil-CoA.
- Ossidazione beta di acidi grassi saturi con un numero dispari di atomi di carbonio
In quegli acidi grassi del numero dispari di atomi di carbonio (che non sono molto abbondanti), la molecola dell'ultimo ciclo di degradazione ha 5 atomi di carbonio, quindi la sua frammentazione produce una molecola di acetil-CoA (che entra nel ciclo di Krebs) e un altro di Propionil-Coa.
Propionil-CoA deve essere carbossilato (reazione dipendente da ATP e bicarbonato) dall'enzima carbossilasi propionil-CoA, che forma un composto noto come d-metilmalonil-CoA, che deve essere epimerizzato nella sua forma "L".
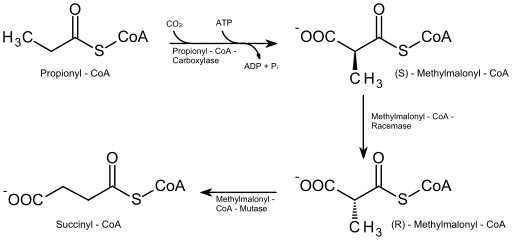 Ossidazione beta di acidi grassi di numeri dispari (Fonte: Eleska [CC0] tramite Wikimedia Commons)
Ossidazione beta di acidi grassi di numeri dispari (Fonte: Eleska [CC0] tramite Wikimedia Commons) Il composto risultante dall'epimerizzazione viene successivamente convertito in succinil-CoA mediante azione dell'enzima L-metilmalonil-CoA mutasi e questa molecola, nonché acetil-CoA, entra nel ciclo di acidi degli agrumi.
- Ossidazione beta di acidi grassi insaturi
Molti lipidi cellulari hanno catene di acidi grassi con insaturazione, cioè hanno uno o più doppi legami tra i loro atomi di carbonio.
L'ossidazione di questi acidi grassi è un po 'diversa da quella degli acidi grassi saturi, poiché due enzimi aggiuntivi, una rabbia isomerasi e 2,4-dieno-coa reduttasi, sono responsabili dell'eliminazione di questa insaturazione in modo che questi acidi grassi possano essere un substrato di substrato l'enzima-coa idratasa.
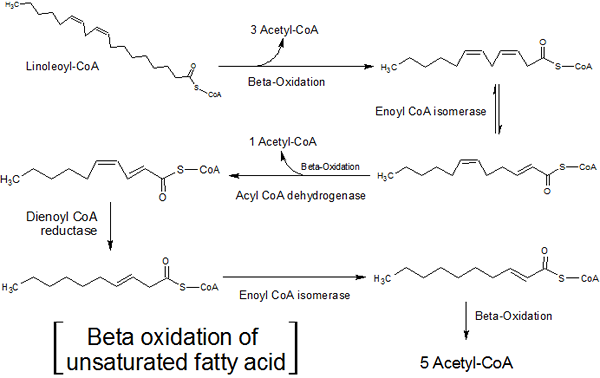 Ossidazione beta di acidi grassi insaturi (fonte: hajime7basketball [CC BY-SA 3.0 (https: // creativeCommons.Org/licenze/by-sa/3.0)] via Wikimedia Commons)
Ossidazione beta di acidi grassi insaturi (fonte: hajime7basketball [CC BY-SA 3.0 (https: // creativeCommons.Org/licenze/by-sa/3.0)] via Wikimedia Commons) La rabbia isomerasi agisce su acidi grassi monoinsaturi (con una singola insaturazione), nel frattempo l'enzima 2,4-dieno-coa reduttasi reagisce con acidi grassi polinsaturi (con due o più insaturazione).
- Beta Extra -Roman Ossidation
L'ossidazione beta degli acidi grassi può anche verificarsi all'interno di altri organelli citosolici come i perossisomi, ad esempio, con la differenza che gli elettroni che vengono trasferiti su FAD+ non vengono consegnati alla catena respiratoria, ma direttamente sull'ossigeni.
Può servirti: alstroemeria: caratteristiche, habitat, cura, specieQuesta reazione produce perossido di idrogeno (l'ossigeno è ridotto), composto che viene eliminato dall'enzima della catlasi, specifico per questi organelli.
Prodotti di ossidazione beta
L'ossidazione degli acidi grassi produce molta più energia del degrado dei carboidrati. Il prodotto principale dell'ossidazione beta è l'acetil-CoA prodotto in ogni fase della porzione ciclica del percorso, tuttavia, altri prodotti sono:
- AMP, H+ e pirofosfato (PPI), prodotto durante l'attivazione.
- FADH2 e NADH, per ogni acetil-CoA prodotto.
- Succinil-CoA, ADP, PI, per acidi a catena dispari.
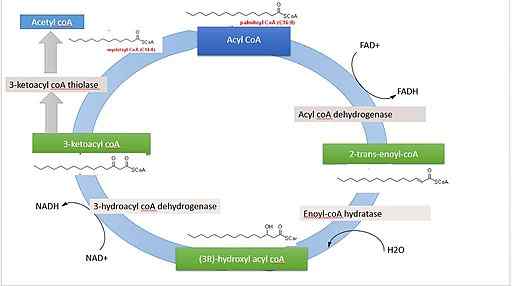 Ossidazione beta dell'acido palmitico (fonte: 'Rojinbkht [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)] via Wikimedia Commons)
Ossidazione beta dell'acido palmitico (fonte: 'Rojinbkht [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)] via Wikimedia Commons) Se consideriamo come esempio la completa ossidazione dell'acido palmitico (palmitato), un acido grasso di 16 atomi di carbonio, la quantità di energia che si verifica è più o meno equivalente a 129 molecole ATP, che provengono dai 7 giri deve completare il ciclo.
Regolamento
La regolazione dell'ossidazione beta degli acidi grassi nella maggior parte delle cellule dipende dalla disponibilità di energia, non solo correlata ai carboidrati ma con gli stessi acidi grassi.
Gli animali controllano la mobilizzazione e, quindi, il degrado dei grassi da parte degli stimoli ormonali, che sono allo stesso tempo controllati da molecole come AMPC, ad esempio.
Nel fegato, il principale organo di degradazione dei grassi, la concentrazione di malonil-CoA è estremamente importante per la regolazione dell'ossidazione beta; Questo è il primo substrato impegnato nella via della biosintesi degli acidi grassi.
Quando il malonil-CoA si accumula in grandi proporzioni, promuove la biosintesi degli acidi grassi e inibisce il trasportatore mitocondriale o la navetta acil-carnitina. Quando la sua concentrazione diminuisce, l'inibizione cessa e l'ossidazione beta viene attivata.
Riferimenti
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biochimica (3 ° ed.). San Francisco, California: Pearson.
- Nelson, d. L., & Cox, m. M. (2009). Principi di biochimica Lechinger. Omega Editions (5 ° ed.).
- Rawn, j. D. (1998). Biochimica. Burlington, Massachusetts: Neil Patterson Publishers.
- Schulz, h. (1991). Ossidazione beta di acidi grassi. Biochimica et Biophysica Acta, 1081, 109-120.
- Schulz, h. (1994). Regolazione dell'ossidazione degli acidi grassi nel cuore. Recensione critica, 165-171.
- Schulz, h., & Kunau, W. (1987). Beta-ossidazione di acidi grassi insaturi: una via di revica. Tibs, 403-406.
- « Circuito produttivo delle fasi principali della carne
- Concetto di sublimazione inversa ed esempi »