

Struttura, funzioni, proprietà, sintesi
- 975
- 19
- Lino Lombardi
Lui Uracile È una base di azoto di pirimidina, che si trova in acido ribonucleico (RNA). Questa è una delle caratteristiche che differenziano l'RNA dall'acido desossiribonucleico (DNA), poiché quest'ultimo ha Timina invece di Uracile. Entrambe le sostanze, Uracile e Timina, differiscono solo che il secondo abbia un gruppo metilico.
Dal punto di vista evolutivo, è stato proposto che l'RNA fosse la prima molecola che immagazzinava informazioni genetiche e lavorava come catalizzatore nelle cellule, prima del DNA e degli enzimi. Per questo motivo, si pensa che Uracile abbia avuto un ruolo chiave nell'evoluzione della vita.
Fonte: Kemikungen [dominio pubblico] Negli esseri viventi, l'uracile non si trova liberamente, ma comunemente nucleotidi monofosfato (UMP), difosfato (UDP) e trfosfato (UTP). Questi nucleotidi di uracile hanno funzioni diverse, come la biosintesi dell'RNA e il glicogeno, l'interconversione isomerica degli zuccheri e la regolazione della sintasi sintasi.
[TOC]
Struttura e proprietà
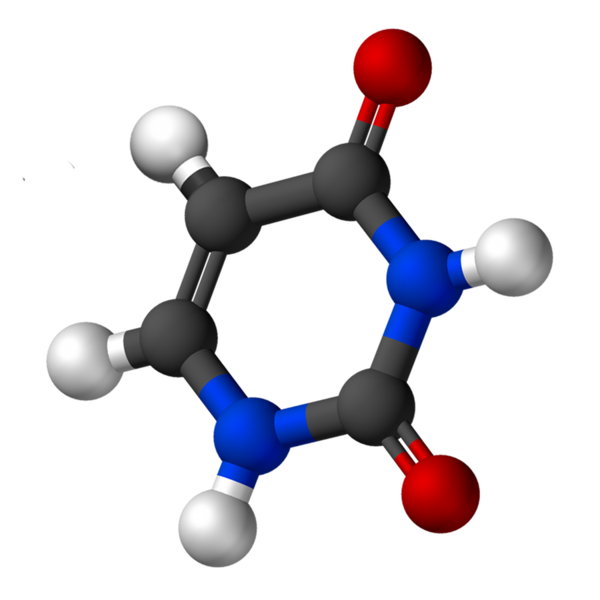
Uracil, chiamato 2.4-diossipiridina, ha la formula empirica C4H4N2O2, il cui peso molecolare è 112,09 g/mol ed è purificato come polvere bianca.
La struttura dell'uridina è un anello eterociclico con quattro atomi di carbonio e due azoto, con doppi legami alternativi. È planare.
Ha una solubilità di 50 mg/ml, 25 ºC, in idrossido di sodio 1 m e un PKA tra 7.9 e 8.2. La lunghezza d'onda in cui si verifica la sua massima assorbanza (ʎMax) È tra 258 e 260 nm.
Biosintesi
Esiste un percorso comune per la biosintesi dei nucleotidi della pirimidina (uracile e citochina). Il primo passo è la biosintesi del fosfato di carbamoil da co2 e NH4+, che è catalizzato da carbamoil sintetasi fosfato.
La pirimidina è costruita in fosfato carboidale e aspartato. Entrambe le sostanze reagiscono e formano N-carbamoilasPartato, Reazioni catalizzate dall'aspartato Trancabamoilasa (Atcasa). La chiusura dell'anello di pirimidina può essere disidratata dalla diidrotasi e produce l-diidrotatao.
La l-diidrotota viene ossidata e trasformata in orotato; L'accettore di elettroni è il NAD+. È una reazione catalizzata dalla diidroorota deidrogenasi. Il passo successivo consiste nel trasferimento del gruppo fosforbosil. Forma gradidilata (WPO) e piroffosfat inorganico (PPI), catalizzato dalla fosforibosil orotato transferasi.
L'ultimo passo consiste nella decarbossilazione dell'anello di gruppo pirimidina (WPO). Forma di Uridilaato (Uridin-5'-Monofosfato, UMP), che è catalizzata da una decarbossilasi.
Quindi, attraverso la partecipazione di una chinasi, un gruppo di fosfato viene trasferito dall'ATP all'UMP, formando UDP (Uridin-5'-difosfato). Quest'ultimo viene ripetuto, formando UTP (Uridin-5'-triffosfato).
Può servirti: rami di biologia e cosa studianoRegolazione della biosintesi
Nei batteri, la regolazione della biosintesi della pirimidina si verifica attraverso un feedback negativo, a livello di aspartato di trancabamoilasa (ATCASA).
Questo enzima è inibito da CTP (Citidin-5'-trofosfato), che è il prodotto finale della via della biosintesi delle pirimidine. Atcas.
Negli animali, la regolazione della biosintesi della pirimidina si verifica attraverso un feedback negativo, a livello di due enzimi: 1) fosfato di carbamoil sintasi II, che è inibita da UTP e attivata da ATP e PRPP; e 2) l'OMP discarbossilasi, che è inibita dal prodotto della reazione che lei stessa catalizza, l'UMP. La velocità di biosintesi dell'OMP varia con la disponibilità di PRPP.
Funzione nella biosintesi dell'RNA
Uracile è presente in tutti i tipi di RNA, come l'RNA messaggero (MNA), l'RNA di trasferimento (ARNT) e l'RNA ribosomiale (RNA). La biosintesi di queste molecole avviene attraverso un processo chiamato trascrizione.
Durante la trascrizione, le informazioni contenute nel DNA vengono copiate nell'RNA attraverso un RNA PON polimerasi. Il processo inverso, in cui le informazioni contenute nell'RNA vengono copiate nel DNA, si verificano in alcuni virus e piante attraverso la trascrittasi inversa.
La biosintesi dell'RNA necessita di nucleosidi Tryposphate (NTP), vale a dire: trifosfato uridina (UTP), citidina tripospha (CTP), adenina adenina adenina (ATP) e guanina trifosfato (GTP). La reazione è:
(RNA)n rifiuti + Ntp -> (RNA)N+1 rifiuti + ppi
L'idrolisi del pirofosfato inorganico (PPI) fornisce energia per la biosintesi dell'RNA.
Funzione nella biosintesi dello zucchero
Gli esteri di zucchero sono molto comuni negli organismi viventi. Alcuni di questi esteri sono le difosfas di esteri nucleosidici, come UDP-Sugar, che sono molto abbondanti nelle cellule. UDP-Sugar coinvolge la biosintesi di disaccaridi, oligosaccaridi e polisaccaridi.
Nelle piante, la biosintesi del saccarosio avviene in due modi: un principale e un secondario.
Il percorso principale consiste nel trasferimento del d-glucosio dall'UDP-D-glucosa al d-frocease per formare saccarosio e UDP. La strada secondaria include due passaggi: inizia con UDP-D-glucosio e fruttosio-6-fosfato e culmina con la formazione di saccarosio e fosfato.
Nelle ghiandole mammarie, la biosintesi del lattosio avviene da UDP-D-galattosio e glucosio.
Può servirti: rami di biochimicaNelle piante, la biosintesi della cellulosa viene effettuata mediante condensazione continua di rifiuti beta-D-glicosil. Allo stesso modo, la biosintesi di amilosio e amilopectina richiede UDP-glucosio come substrato di donatore di glucosio alla catena di crescita.
Negli animali, sia UDP-glucosio che ADP-glucosio sono usati per la biosintesi del glicogeno. Allo stesso modo, la biosintesi del solfato di condroitina richiede UDP-xilosio, UDP-galattosio e UDP-glucuronato.
Funzione nell'interconversione isomerica degli zuccheri
La conversione del galattosio in un intermediario di glicolisi avviene attraverso la strada di Leloir. Uno dei passaggi di questo percorso è catalizzato dall'enzima UDP-galattosio-4-epimerasi, che facilita l'interconversione di UDP-galattosio a UDP-glucosio.
Funzione nella biosintesi delle glicoproteine
Durante la biosintesi delle glicoproteine, le proteine attraversano i sacchi cis, medi e trans dell'apparato Golgi.
Ognuna di queste borse ha una serie di enzimi che elaborano glicoproteine. Monomeri di zucchero, come glucosio e galattosio.
Gli esosi nucleotidi vengono trasportati in serbatoi di Golgi da Antiporte. L'UDP-galattosio (UDP-GAL) e l'UDP-N-Acetilgalattosamina (UDP-GalNAC) entrano nei serbatoi dal citosol mediante scambio da UMP.
Nel serbatoio del Golgi, un fosfatasi idrolizza un gruppo fosfato della forma UDP e UMP e PI. L'UDP proviene dalle reazioni catalizzate da galattosiltransferasi e N-acetilgalattosamiltransferasi. L'UMP formata dalla fosfatasi serve a scambiare i nucleotidi-esosio.
Funzione nella regolazione della sintasi
Un meccanismo per la regolazione della glutammina sintasi è la modifica covalente, che consiste in adenilazione, che inattiva e fianco, che la attiva. Questa modifica covalente è reversibile e catalizzata dall'adeniltransferasi.
L'attività dell'adeniltransferasi è modulata dall'unione della proteina PII, che è regolata da una modifica covalente, uridinilazione.
Sia l'uridililazione che l'offset sono eseguiti dall'uridililtransferasi. In questo enzima, l'attività di uridilazione è dovuta a glutammina e fosfato ed è attivata dall'unione di alfa-zotoglutarato e ATP al PII.
Funzione nell'edizione RNA
Alcuni RNM vengono modificati prima della traduzione. In alcuni organismi eucariotici, come Trypanosoma Brucei, C'è un'edizione dell'RNA della trascrizione del gene subunità II del citocromo ossidasi. Ciò accade inserendo i rifiuti di uracile, una reazione catalizzata dall'uridiltransferasi terminale.
Può servirti: ricciolo: composizione, parti, importanzaUn RNA guida, complementare al prodotto modificato, agisce come temperato per il processo di editing. Le coppie di basi formate tra la trascrizione iniziale e la guida RNA implica coppie di basi g = u che non sono record watson e sono comuni nell'RNA.
Biosintesi UDP-glucosio
In condizioni fisiologiche, la biosintesi del glicogeno dal glucosio-1-fosfato è termodinamicamente impossibile (ΔG positivo). Per questo motivo, prima della biosintesi, si verifica l'attivazione del glucosio-1-fosfato (G1P). Questo G1P e UTP combinano la reazione per formare il glucosio difosfato uridina (UDP-glucosio o UDPG).
La reazione è catalizzata dalla pirofosforillasi dell'UDP-glucosio ed è la seguente:
G1P + UTP -> UDP -GLUCOSA + 2PI.
La variazione di energia libera di Gibbs in questo passaggio è grande e negativa (-33,5 kJ/mol). Durante la reazione di ossigeno, il G1P attacca l'atomo di alfo alfa fosforo del pirofosfato UDP-glucosio e inorganico (PPI). Quindi, il PPI viene idrolizzato da un pirofosfati inorganici, la cui energia di idrolisi è quella che guida la reazione generale.
UDP-Glucosio è una sostanza "ad alta energia". Permette di formare i legami glicosidici tra il residuo di glucosio e la crescente catena di polisaccaride. Questo stesso principio energetico è applicabile alle reazioni in cui partecipa l'UDP-Sugar, come la biosintesi di disaccaridi, oligosaccaridi e glicoproteine.
Uracil DNA Glycosilasa
Ci sono lesioni del DNA che si verificano spontaneamente. Una di queste lesioni è lo spronta. In questo caso, la riparazione avviene a causa della base di DNA modificata per mezzo di un enzima chiamato DNA Uracil glicosilasa.
L'enzima Uracil DNA Glycosilasa elimina la citochina danneggiata (uracile), producendo un residuo di desossiribosio che manca della base di azoto, chiamato sito AP (sito apurinico-aprimidinico).
Quindi, l'enzima AP endonucleasi fa un taglio nello scheletro fosfodiestro del sito AP, eliminando il residuo di zucchero-fosfato. Il DNA polimerasi I ripristina il filo danneggiato.
Riferimenti
- Bohinski, r. 1991. Biochimica. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biochimica. Redazione editoriale, Barcellona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, p., Baltimora, d., Darnell, J. 2003. Biologia cellulare e molecolare. Editoriale Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Messico, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Leynninger-Principles of Biochemistry. W.H. Freeman, New York.
- Voet, d. e Voet, J. 2004. Biochimica. John Wiley and Sons, USA.
- « Formula di costo marginale, calcolo ed esercizi
- Caratteristiche, struttura, funzioni delle pirimidine »