

Teorie principali del patrimonio postmendeliano

- 2203
- 102
- Silvano Montanari
IL Postmendelian Heritage Include tutte le idee e/o le opere che sono sorte dopo gli studi pubblicati dal naturalista religioso Gregorio Mendel nel 1865 e che spiegano i fenomeni o comportamenti genetici diversi da quelli descritti da questo.
Gregorio Mendel è considerato il padre della genetica, da quando ha studiato le croci tra le piante di pisello, ha stabilito le prime leggi dell'eredità più di 100 anni fa. La prima legge, chiamata "Legge della segregazione", propone che i geni possano esistere in forme diverse o alternative.
 Gregorio Mendel (fonte: autore sconosciuto / dominio pubblico, via Wikimedia Commons)
Gregorio Mendel (fonte: autore sconosciuto / dominio pubblico, via Wikimedia Commons) Queste forme alternative sono chiamate "alleli" e influenzano direttamente i caratteri fenotipici di una specie. Gli alleli possono determinare il colore dei capelli, degli occhi, del tipo di capelli, ecc.
Nella sua prima legge, Mendel ha proposto che ogni caratteristica fenotipica trasmessa dai genitori ai propri figli sia controllato da due alleli, ciascuno da un genitore (una delle "madre" e un'altra del "padre").
Secondo questo, quando i genitori forniscono alleli diversi, la progenie risulta con una miscela, in cui uno degli alleli è dominante (quello che è espresso nel fenotipo) e l'altro è recessivo (non si vede o è "latente ").
La seconda legge di Mendel è conosciuta come "Legge dell'eredità indipendente", poiché nel suo Cruces Mendel ha osservato che il modello di eredità che caratterizzava gli alleli che definivano una caratteristica non influenzava il modello di eredità degli alleli definiti.
[TOC]
Teorie ereditarie postmendeliane
Poiché Gregor Mendel ha proposto le sue famose leggi più di 100 anni nuovi meccanismi sono stati chiariti per spiegare l'eredità dei personaggi, molti dei quali si potrebbero dire che contraddicono o differiscono considerevolmente da quelli descritti da Mendel.
Esempio dei fenomeni descritti dalla scienza "moderna", i cui progressi sono stati possibili solo grazie al progresso delle strategie sperimentali e dei metodi di ricerca, sono l'eredità legata al sesso, al dominio incompleto e al codice, tra gli altri.
- Eredità legata al sesso
L'eredità legata al sesso ha a che fare con i geni trovati solo nei cromosomi sessuali. Nell'uomo, il sesso è determinato dai cromosomi X e Y. Le donne hanno due copie di x (xx) e gli uomini uno di x e un altro di y (xy).
Può servirti: geni collegati Eredità collegata al materno via (fonte: Jordi Picart/CC BY-SA (https: // creativeCommons.Org/licenze/by-sa/4.0) via Wikimedia Commons)
Eredità collegata al materno via (fonte: Jordi Picart/CC BY-SA (https: // creativeCommons.Org/licenze/by-sa/4.0) via Wikimedia Commons) I geni trovati in ciascuno di questi due cromosomi sono quelli che contribuiscono a quei tratti che sono collegati al sesso. I geni sul cromosoma X sono presenti negli uomini e nelle donne, ma i geni di e sono solo negli uomini.
L'eredità legata al sesso è stata osservata e descritta per la prima.
Bateson ha realizzato croci sperimentali di polli con piumaggi di diversi colori, alcuni con piume scure e lisce e altri con piume lisce e con un motivo a strisce.
Attraverso queste croci osservò che, nella prole, i maschi risultanti avevano sempre piumaggi con un motivo a strisce.
Solo in terza generazione, quando un maschio è graffiato con una piume scura e liscia incrociate, sono stati ottenuti alcuni galli con piumaggio nero. Questa proporzione derivante dalle croci ha sfidato le proporzioni proposte da Mendel.
Eredità collegata a x
Dopo il lavoro di Bateson, Thomas Henry Morgan ha trovato una spiegazione logica per risultati così strani ottenuti da Bateson, attribuendo la differenza di colorazione nei polli a cui il motivo a strisce era codificato da un allele dominante che si trovava sul cromosoma X.
Nei polli, a differenza dell'essere umano, il sesso eterogametico (XY) corrisponde alle femmine, nel frattempo il sesso omogamico corrisponde ai maschi (xx).
L'allele studiato da Bateson e Morgan, quindi, è stato espresso da tutti quei maschi con un piumaggio a strisce, ma quando un maschio ha ereditato due cromosomi X con l'allele corrispondente alla colorazione nera, allora sono apparsi uomini di piumaggio scuro e senza motivi graffiati.
- Dominanza incompleta
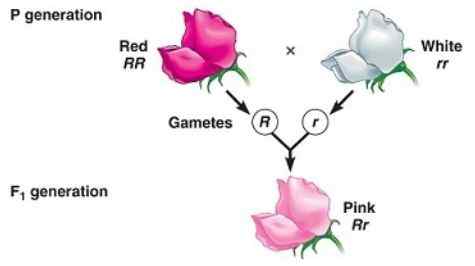 Fenotipo intermedio nella generazione F1 causata da un dominio incompleto
Fenotipo intermedio nella generazione F1 causata da un dominio incompleto Il dominio incompleto è quando il fenotipo osservato nella prole che deriva dall'incrocio di due individui omozigoti (uno dominante e un altro recessivo) è un fenotipo "intermedio" tra il dominante e il recessivo.
Ha a che fare con l '"incapacità" dell'allele "dominante" di mascherare completamente l'espressione dell'allele recessivo, motivo per cui il fenotipo risultante corrisponde a una miscela delle caratteristiche codificate da entrambi gli alleli, spesso descritto come un "fenotipo diluito "Dal dominante.
Può servirti: monosomieAd esempio, nella notte -Year Plants (Mirabilis Jalapa) Il colore tipico dei petali di fiori selvatici è rosso. Quando si ottiene una pianta con petali rossi con una pianta di linea pura con petali bianchi, si ottiene che tutta la progenie è petali rosa (rosa).
- Codominanza
Quando due individui omozigoti dominanti vengono attraversati e nella prole c'è un terzo fenotipo che corrisponde alle due caratteristiche determinate dai due alleli dei genitori, parliamo di codominanza.
In questo caso, seguendo un esempio simile a quello dei fiori dell'esempio precedente, il "terzo fenotipo" non corrisponderebbe ai fiori rosa, ma a una pianta con fiori bianchi e fiori rossi nello stesso stelo, poiché i due alleli " co -cake ”, e nessuno maschera l'altro.
Molte volte, tuttavia, il codice incompleto e il dominio sono difficili da distinguere fenotipicamente, poiché entrambi producono il fenotipo di un individuo che sembra esprimere due alleli diversi.
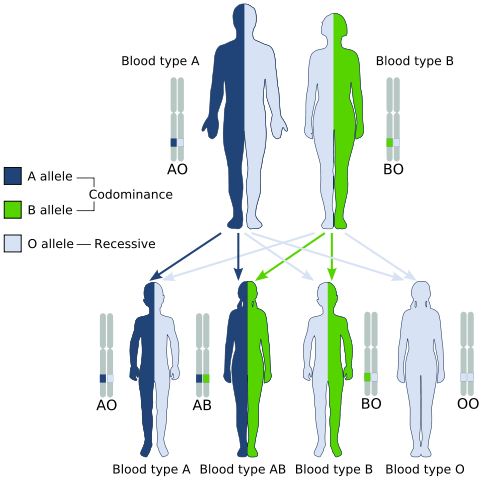 Codominance, esempio di un tipo di eredità postmendeliana (Fonte: Gyssinemrabettalk✉ Questa immagine W3C-Runspecified è stata creata con Inkcape. / Dominio pubblico, via Wikimedia Commons)
Codominance, esempio di un tipo di eredità postmendeliana (Fonte: Gyssinemrabettalk✉ Questa immagine W3C-Runspecified è stata creata con Inkcape. / Dominio pubblico, via Wikimedia Commons) Tuttavia, genotipicamente, gli individui presentano.
Un classico esempio di codominanza è il tipo di gruppo sanguigno nell'uomo, poiché gli alleli che codano per gli antigeni A e B nelle popolazioni umane sono codominanti.
Cioè, quando gli alleli A e B si trovano insieme nelle cellule di un individuo, ha un tipo di sangue di sangue.
- Alleli letali
Sono alleli recessivi per un po 'di carattere che, quando sono omozigoti, sono letali per l'individuo. Gli individui che presentano questi alleli non nascono, poiché evitano il normale funzionamento dei macchinari molecolari ed evitano la traduzione dei geni.
- Interazioni geniche
Molti posti nel genoma di un individuo interagiscono con altre parti dello stesso genoma, cioè che la modifica puntuale di un allele all'interno del genoma può condizionare direttamente l'espressione di altri alleli all'interno delle cellule di un individuo.
Può servirti: impatto sociale, economico e ambientale dell'ingegneria geneticaQueste interazioni sono difficili da determinare e sono classificate in due diversi tipi: i geni che interagiscono sulla stessa via genetica e quelli che lo fanno su percorsi genetici diversi.
Interazioni su diverse rotte genetiche
Un esempio classico e noto di geni che interagiscono in diverse rotte genetiche ha a che fare con i geni che determinano il colore della pelle dei serpenti di mais (PANTHEROPHIS GUTTATUS), il cui colore comune è un motivo ripetitivo di punto nero e arancione.
 Corn Snake (Fonte: Factumquintus, via Wikimedia Commons)
Corn Snake (Fonte: Factumquintus, via Wikimedia Commons) Questo fenotipo è prodotto da due diversi pigmenti che sono sotto stretto controllo genetico. Un gene determina l'arancia e questo ha due alleli: uno o+, che codifica per il pigmento e un altro o-, che determina l'assenza totale del pigmento arancione.
Un altro gene diverso determina la presenza o l'assenza del pigmento nero, gli alleli di questo gene sono noti come B+, quello che determina la presenza del pigmento nero e B-, che determina l'assenza del pigmento.
Entrambi i geni non sono collegati e determinati indipendentemente dal colore dei serpenti, cioè un serpente con il genotipo o+/b+ è un serpente normale con fenotipo arancione e nero, ma qualsiasi variazione genotipica comporterà un fenotipo diverso.
Interazioni sulla stessa via genetica
Il classico esempio di questi geni è ciò che è stato descritto per i fiori di "campane" (dalla famiglia Campanulae), il cui fenotipo "selvaggio" corrisponde alle piante con fiori blu.
Il colore blu dei fiori di campana non è determinato da un gene, ma da una via metabolica in cui diversi enzimi, ciascuno codificati da un gene diverso, partecipano alla sintesi di un pigmento.
Pertanto, ci sono fenotipi caratterizzati dall'essere il risultato dell'interazione di diversi geni, i cui prodotti partecipano alla stessa via metabolica e in cui la mutazione o la cattiva funzione di una di queste, produce un fenotipo, in questo caso, di fiori bianchi, depigmentato.
Riferimenti
- Ceccalli, d. (2019). Storia dell'eredità.
- Griffiths, p., & Stotz, K. (2013). Genetica e filosofia: un'introduzione. Cambridge University Press.
- Lewin, b., Krebs, j. E., Goldstein, e. S., & Kilpatrick, s. T. (2014). Geni XI di Lewin. Jones & Bartlett Publishers.
- McCleran, g. E., Vogler, g. P., & Plomin, r. (millenovecentonovantasei). Genetica e medicina comportamentale. Medicina comportamentale, 22(3), 93-102.
- Richmond, m. L. (2008). Il programma di ricerca pre-e post-mendeliano di William Bateson in "Ereditarietà e sviluppo.". Una storia culturale di ereditarietà IV: ereditarietà nel secolo del gene, 343, 213-242.
- « Principi di irritabilità cellulare, componenti, risposte (esempi)
- Megafauna estinto e attuale (con esempi) »