

Imballaggio DNA
- 3994
- 208
- Enzo De Angelis
Schema di imballaggio del DNA. Fonte: Thomas Splettstoesser (www.Scistilo.com), cc di 4.0, Wikimedia Commons Cos'è l'imballaggio DNA?
Lui Imballaggio DNA È un termine che definisce la compattazione controllata del DNA all'interno cellulare. Il DNA è una molecola estremamente lunga che, inoltre, interagisce sempre con un'enorme varietà di diverse proteine (nucleoproteine). Con loro si forma la cromatina, che è la sostanza che compone i cromosomi.
Per l'elaborazione, l'eredità e il controllo dell'espressione dei geni che il DNA adotta una particolare organizzazione spaziale. La cellula raggiunge il controllo rigoroso del DNA a diversi livelli di compattazione.
I virus hanno strategie di imballaggio diverse dei loro acidi nucleici. Uno dei preferiti è la formazione di spirali compatte. Si potrebbe dire che i virus sono acidi nucleici confezionati nelle proteine che li coprono, proteggono e mobilitano.
Nei procarioti, il DNA è associato a proteine che determinano la formazione di legami complessi in una struttura chiamata nucleoide. Il livello massimo di compattazione del DNA in una cellula eucariotica, d'altra parte, è il cromosoma mitotico o meiotico.
Struttura del DNA
Il DNA è costituito da due bande antipaallali (corrono nella direzione opposta) che formano una doppia elica. Ognuno di essi presenta uno scheletro di legami di fosfodié su cui sono uniti gli zuccheri legati alle basi di azoto.
All'interno della molecola, le basi di azoto di una banda formano ponti idrogeno (due o tre) con la banda complementare.
In una molecola come questa, gli angoli di collegamento più importanti mostrano una rotazione libera. I legami di base della sughero azoto, il fosfato del gruppo di zucchero e il legame con fosfoditester sono flessibili.
Ciò consente al DNA, visto come un'asta flessibile, mostra una certa capacità di piegarsi e rotolare. Questa flessibilità consente di adottare strutture locali complesse e formare legami di interazione corta, media e lunga distanza.
Può servirti: qual è la teoria cromosomica dell'eredità? (Sutton e Morgan)Questa flessibilità spiega anche come 2 metri di DNA possono essere mantenuti in ogni cellula diploide di un essere umano. In un gamete (cellula aploide), sarebbe un metro di DNA.
Il nucleoide batterico
Sebbene non sia una regola incrollabile, il cromosoma batterico esiste come una singola molecola di DNA a doppia banda.
La doppia elica è più su se stessa (più di 10 bp per ritorno) producendo così una certa compattazione. Anche i nodi locali sono generati grazie a manipolazioni che sono controllate enzimaticamente.
Inoltre, ci sono sequenze di DNA che consentono ai domini di formarsi in grandi legami. La struttura risultante dal super -collasso e dai legami ordinati è chiamata nucleoide.
Questi sperimentano cambiamenti dinamici grazie ad alcune proteine che forniscono una certa stabilità strutturale al cromosoma compatto. Il grado di compattazione nei batteri e negli archi è così efficiente che potrebbe esserci più di un cromosoma nucleoide.
Il DNA nucleoide compatto procarioti almeno circa 1.000 volte. La struttura topologica nucleoide è una parte fondamentale della regolazione dei geni che il cromosoma trasporta. Cioè la struttura e la funzione costituiscono la stessa unità.
Livelli di compattazione cromosomica eucariotica
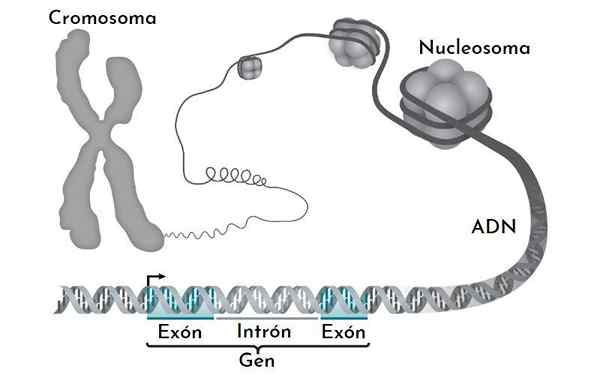
Il DNA nel nucleo eucariotico non è nudo. Interagisci con molte proteine, la più importante delle quali sono gli istoni. Le istoni sono piccole proteine caricate positivamente che si legano al DNA in modo specifico.
Nel nucleo, ciò che osserviamo è un DNA complesso: gli istoni, che chiamiamo cromatina. La cromatina altamente condensata, che generalmente non è espressa, è eterocromatina. Al contrario, il meno compatto (più aperto), o euchromatina, è la cromatina con geni che sono espressi.
Può servirti: pura lineaLa cromatina ha diversi livelli di compattazione. Il più elementare è il nucleosoma. Sono seguiti dalle fasce di fibra del solenoide e dalle cromatina di interfaccia. Solo quando un cromosoma è diviso è che i livelli di compattazione massimi sono mostrati.
Il nucleosoma
Il nucleosoma è l'unità di base dell'organizzazione della cromatina. Ogni nucleosoma è formato da un octa di istoni che formano una specie di tamburo.
L'ottamero è formato da due copie di ciascuno degli istoni H2A, H2B, H3 e H4. Intorno a loro, il DNA dà quasi 1.7 turni. È seguito da una frazione di DNA libera chiamata Linker, 20 bp, associata all'istone H1 e poi a un altro nucleosoma. La quantità di DNA in un nucleosoma e quella che si unisce è circa 166 coppie di basi.
Questa fase di imballaggio del DNA compatto la molecola circa 7 volte. Cioè, passiamo un metro a poco più di 14 cm di DNA.
Questo packaging è possibile perché gli istoni positivi annullano il carico negativo del DNA e la conseguente autorepulsione elettrostatica. L'altro motivo è che il DNA può essere piegato in modo tale da poter girare l'ottamero dell'istone.
Fibra da 30 nm
I conti in una collana che formano molti nucleosomi successivi sono inoltre iscritti a una struttura più compatta.
Anche se non siamo sicuri di quale struttura adotta davvero, sappiamo che raggiunge uno spessore di circa 30 nm. Questa è la fibra così chiamata da 30 nm e per la sua formazione e stabilità l'istone H1 è fondamentale.
La fibra da 30 nm è l'unità strutturale di base dell'eterocromatina. Quello dei nucleosomi lassisti, quello di euchromatina.
Legami e curve
La fibra da 30 nm, tuttavia, non è completamente lineare. Al contrario, forma legami lunghi circa 300 nm, in modo tortuoso, su una matrice proteica poco conosciuta.
Può servirti: genetica forense: storia, oggetto di studio, metodologiaQuesti legami su una matrice proteica formano una fibra di cromatina più compatta di 250 nm di diametro. Infine, si allineano come una semplice elica spessa 700 nm, dando origine a uno dei cromatidi sorelle di un cromosoma mitotico.
Alla fine, il DNA nella cromatina nucleare compatta circa 10.000 volte sul cromosoma cellulare in divisione. Nel core dell'interfaccia la sua compattazione è anche alta, poiché è circa 1.000 volte rispetto al DNA "lineare".
La compattazione meiotica del DNA
Nel mondo della biologia dello sviluppo si dice che la gameteogenesi resetta l'epigenoma. Cioè, cancella i marchi del DNA che la vita di coloro che hanno dato origine a Gameto hanno prodotto o sperimentato.
Questi marchi includono la metilazione del DNA e le modifiche covalenti degli istoni (codice dell'istone). Ma non tutto l'epigenoma viene ripristinato. Ciò che rimane con i marchi sarà responsabile dell'impronta genetica paterna o materna.
Il ripristino implicito alla gametoogenesi è più facile vederlo nello sperma. Nello sperma, il DNA non è confezionato con istoni. Pertanto, le informazioni associate alle sue modifiche nel corpo produttore, in generale, non sono ereditate.
In sperma, il DNA è confezionato grazie all'interazione con le proteine dell'Unione non specifiche per il DNA, chiamate protamine. Queste proteine formano ponti disolfuro tra loro, contribuendo così alla formazione di strati traboccanti che non si respingono elettrostaticamente.
Riferimenti
- Packaging del DNA: nucleosomi e cromatina. Estratto dalla natura.com.