

Struttura degli acidi grassi, tipi, funzioni, biosintesi
- 4972
- 773
- Lidia Valentini
IL acidi grassi Sono macromolecole organiche derivate da idrocarburi, che sono composte da lunghe catene di atomi di carbonio e idrogeno che sono idrofobici (sono solubili in grasso) e sono le basi strutturali di grassi e lipidi.
Sono molecole molto diverse che si distinguono l'una dall'altra per la lunghezza delle loro catene di idrocarburi e la presenza, il numero, la posizione e/o la configurazione dei loro doppi legami.
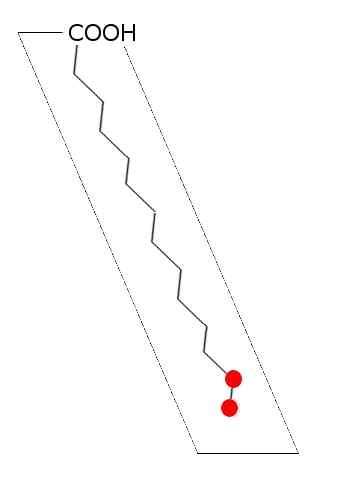
 Schema generale di acido grasso saturo (fonte: laghi.L [CC BY-SA 3.0 (http: // creativeCommons.Org/licenze/by-sa/3.0/)] via Wikimedia Commons)
Schema generale di acido grasso saturo (fonte: laghi.L [CC BY-SA 3.0 (http: // creativeCommons.Org/licenze/by-sa/3.0/)] via Wikimedia Commons) Nei lipidi di animali, piante, funghi e microrganismi come batteri e lieviti, sono stati descritti più di 100 tipi di diversi acidi grassi e sono considerati specie e tessuti specifici nella maggior parte degli esseri viventi.
Gli oli e i grassi che l'uomo consumano quotidianamente sono di origine animale o vegetale, sono principalmente composti da acidi grassi.
[TOC]
introduzione
 Il burro, tra le altre cose, è essenzialmente composto da acidi grassi (fonte: Africa Studio, tramite Pixabay.com)
Il burro, tra le altre cose, è essenzialmente composto da acidi grassi (fonte: Africa Studio, tramite Pixabay.com) Le molecole di acidi grassi svolgono funzioni importanti a livello cellulare, il che li rende componenti essenziali e, poiché alcuni di essi non possono essere sintetizzati dagli animali, devono ottenerle dalla dieta.
Gli acidi grassi sono rari come specie libere nel citosol cellulare, quindi fanno generalmente parte di altri coniugati molecolari come:
- Lipidi, nelle membrane biologiche.
- Trigliceridi o esteri di acidi grassi, che fungono da riserva nelle piante e negli animali.
- Cere, che sono esteri solidi di acidi grassi a catena lunga e alcoli.
- Altre sostanze simili.
Negli animali, gli acidi grassi sono immagazzinati nel citoplasma delle cellule come piccole gocce di grasso composte da un complesso chiamato triacilglicerolo, che non è altro che una molecola di glicerolo a cui si è unita, in ciascuno dei suoi atomi di carbonio, una catena di acidi grassi da legami di tipo estere.
Mentre i batteri hanno acidi grassi corti e solitamente monoinsaturi, in natura è comune.
Struttura
Gli acidi grassi sono molecole anfipatiche, cioè hanno due regioni definite chimicamente: una regione polare idrofila e una regione apolare idrofobica.
La regione idrofobica è costituita da una lunga catena di idrocarburi che, in termini chimici, non è molto reattiva. La regione idrofila, d'altra parte, è composta da un terminale di gruppo carbossilico (-coh), che si comporta come un acido.
Questo gruppo carbossilico terminale o acido carbossilico è ionizzato in soluzione, è altamente reattivo (chimicamente parlando) ed è molto idrofilo, quindi rappresenta un sito di giunzione covalente tra acido grasso e altre molecole.
La lunghezza delle catene di idrocarburi di acidi grassi di solito ha un numero uniforme di atomi di carbonio, e questo è strettamente correlato al processo biosintetico attraverso il quale vengono prodotti, poiché la loro crescita avviene dalle coppie di carboni.
Gli acidi grassi più comuni hanno catene tra 16 e 18 atomi di carbonio e animali, queste catene non sono ramificate.
Classificazione
Gli acidi grassi sono classificati in due grandi gruppi in base alla natura dei collegamenti che li compongono, cioè in base alla presenza di semplici legami o doppi legami tra gli atomi di carbonio delle loro catene di idrocarburi.
Pertanto, ci sono acidi grassi saturi e insaturi.
- Gli acidi grassi saturi hanno solo semplici legami di carbonio - carbonio e tutti i loro atomi di carbonio sono "saturi" o collegati alle molecole di idrogeno.
- Gli acidi grassi insaturi hanno uno o più legami di carbonio a doppio carbonio e non tutti questi sono attaccati a un atomo di idrogeno.
Anche gli acidi grassi insaturi sono divisi in base al numero di insaturazione (doppi legami) in monoinsaturati, quelli con un singolo doppio legame e polinsaturi, che ne hanno più di uno.
Acidi grassi saturi
Di solito hanno tra 4 e 26 atomi di carbonio uniti da semplici collegamenti. Il suo punto di fusione è direttamente proporzionale alla lunghezza della catena, cioè al suo peso molecolare.
Gli acidi grassi che hanno tra 4 e 8 carboni sono liquidi a 25 ° C e sono quelli che compongono oli commestibili, mentre quelli con più di 10 atomi di carbonio sono solidi.
Tra i più comuni è l'acido laurico, che è abbondante nei semi di palma e cocco; L'acido palmitico, che si trova in palma, cacao e burro di maiale e acido stearico, che è in cacao e oli idrogenati.
Sono acidi grassi con molta più stabilità degli acidi grassi insaturi, specialmente di fronte all'ossidazione, almeno in condizioni fisiologiche.
Grazie al fatto che il semplice carbonio - i legami di carbonio possono ruotare.
Acidi grassi insaturi
Questi acidi grassi sono altamente reattivi e inclini alla saturazione e all'ossidazione. Sono comuni nelle piante e negli organismi marini. Coloro che hanno un singolo doppio legame sono conosciuti come monoinsaturi o monoenici, mentre quelli che possiedono più di due sono noti come polienici o polinsaturi.
Può servirti: DNA polimerasiLa presenza di doppi legami è comune tra gli atomi di carbonio tra le posizioni 9 e 10, ma ciò non significa che gli acidi grassi monouinsaturi non si trovano con insaturazione in un'altra posizione.
A differenza degli acidi grassi saturi e insaturi sono elencati non dal gruppo carbossilico terminale, ma in base alla posizione del primo doppio legame C - C. Pertanto, questi sono divisi in due gruppi, omega-6 o ω6 e omega-3 o ω3.
Gli acidi omega-6 hanno il primo doppio legame negli acidi numero 6 e omega-3 e lo hanno in carbonio numero 3. La denominazione ω è data dal doppio legame più vicino al gruppo metilico finale.
I doppi collegamenti possono anche essere trovati in due configurazioni geometriche note come "Cis " E "trans".
La maggior parte degli acidi grassi insaturi naturali hanno configurazione "Cis"E i doppi legami di acidi grassi presenti nei grassi commerciali (idrogenati) si trovano in "trans".
Negli acidi grassi polinsaturi, due doppi legami sono generalmente separati l'uno dall'altro almeno da un gruppo metilico, ovvero un atomo di carbonio attaccato a due atomi di idrogeno.
Funzioni
Gli acidi grassi hanno molteplici funzioni negli organismi viventi e, come menzionato sopra, una delle loro funzioni essenziali come parte essenziale dei lipidi, che sono i componenti principali delle membrane biologiche e una delle tre biomolecole più abbondanti negli organismi vivi in congiunzione proteine e carboidrati.
Sono anche eccellenti substrati energetici grazie a cui sono ottenute grandi quantità di energia sotto forma di ATP e altri metaboliti intermediari.
Alla luce del fatto che gli animali, ad esempio, non sono in grado di conservare carboidrati, gli acidi grassi rappresentano la principale fonte di accumulo di energia che deriva dall'ossidazione degli zuccheri in eccesso.
Gli acidi grassi saturi a catena corta nel colon partecipano alla stimolazione dell'assorbimento d'acqua e al sodio, al cloruro e agli ioni bicarbonato; Inoltre, hanno funzioni nella produzione di muco, nella proliferazione di colonociti (cellule del colon), ecc.
Gli acidi grassi insaturi sono particolarmente abbondanti negli oli commestibili di origine vegetale, che sono importanti nella dieta di tutti gli esseri umani.
 Gli oli che consumiamo quotidianamente sono acidi grassi (fonte: Stevepb, tramite Pixabay.com)
Gli oli che consumiamo quotidianamente sono acidi grassi (fonte: Stevepb, tramite Pixabay.com) Altri partecipano come ligandi di alcune proteine con attività enzimatiche, quindi sono importanti rispetto ai loro effetti sul metabolismo energetico delle cellule in cui si trovano.
Biosintesi
La degradazione degli acidi grassi è noto come β-ossidazione e si verifica nei mitocondri delle cellule eucariotiche. La biosintesi, al contrario, si verifica nel citosol delle cellule animali e nei cloroplasti (organelli fotosintetici) delle cellule vegetali.
È un processo acetil-CoA, malonil-CoA e NADPH, si verifica in tutti gli organismi viventi e in animali "superiori" come i mammiferi. Ad esempio, è molto importante nei tessuti epatici e adiposi, nonché nelle ghiandole mammarie.
Il NADPH utilizzato per questo percorso è principalmente un prodotto delle reazioni di ossidazione NADP-dipendenti della via del fosfato pentosio ossidazione di acidi grassi.
La via della biosintesi, come l'ossidazione β, è altamente regolata in tutte le cellule da effettori areestrici e modificazioni covalenti degli enzimi che partecipano alla regolazione.
-Sintesi malonil-CoA
Il percorso inizia con la formazione di un intermediario metabolico noto come malonil-CoA da una molecola di acetil-CoA ed è catalizzato da un enzima multifunzionale chiamato acetil-CoA carbossilasa.
Questa reazione è una reazione all'aggiunta a una molecola carbossilica (-COH, carbossilazione) dipendente dalla biotina e si verifica in due fasi:
- In primo luogo, il trasferimento, dipendente dall'ATP, di un carbossilico derivato dal bicarbonato (HCO3-) a una molecola di biotina che si trova come un gruppo protesico (non proteina) associato ad acetil-CoA carboxlasi.
- Successivamente, la CO2 viene trasferita in acetil-CoA e viene prodotto Malonil-CoA.
-Reazioni di percorso
Negli animali, la formazione di catene di carboidrati di acidi grassi si verifica ulteriormente attraverso reazioni di condensazione sequenziale catalizzate da un enzima multi -critico e multifunzionale noto come acido grasso sintasi.
Questo enzima catalizza la condensazione di un'unità acetil-CoA e molecole multonil-CoA che sono prodotte dalla reazione della carbossilasi acetil-CoA, durante il quale viene rilasciata una molecola di CO2 per ogni malonil-CoA che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che si aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che si aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che si aggiunge che aggiunge che si aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che siroro in via di malonio-Couma che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che siroro in via di malonica-Couma che aggiunge che aggiunge che aggiunge che aggiunge che aggiunge che siroro.
Gli acidi grassi in crescita sono sterilizzati da una proteina chiamata "proteina portante acilica" o ACP, che forma tioé con gruppi acilici. In E. coli Questa proteina è un polipeptide da 10 kDa, ma negli animali fa parte del complesso acido grasso sintasi.
Può servirti: noxaLa rottura di questi legami Tioéster rilascia grandi quantità di energia, il che lo rende possibile, termodinamicamente, il verificarsi delle fasi di condensa sulla via biosintetica.
Complesso di acido grasso sintasi
Nei batteri, l'attività dell'acido sintasi corrisponde effettivamente a sei enzimi indipendenti che usano acetil-CoA e malonil-CoA per formare acidi grassi e con cui sono associate sei diverse attività enzimatiche.
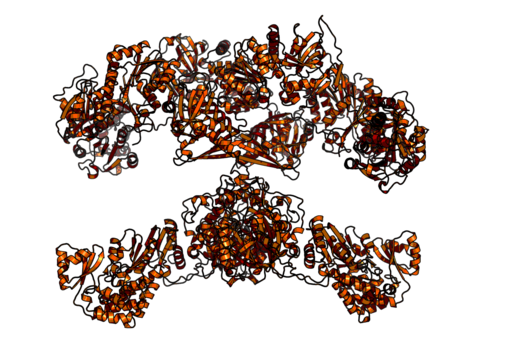 Acido grasso complesso omodimerico e multifunzionale sintasi di animali (fonte: Boehringer Ingelheim [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)] via Wikimedia Commons)
Acido grasso complesso omodimerico e multifunzionale sintasi di animali (fonte: Boehringer Ingelheim [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)] via Wikimedia Commons) Nei mammiferi, d'altra parte, l'acido grasso sintasi è un complesso enzimatico omodimico multifunzionale di circa 500 kDa di peso molecolare, che ha sei diverse attività catalitiche e con le quali è associata la proteina portante acida.
Passaggio 1: reazione abilmente
I gruppi di Tiol nei rifiuti di cisteina responsabili dell'Unione degli intermediari metabolici nell'enzima ACP devono essere caricati, prima dell'inizio della sintesi, con i gruppi acidi necessari.
Per fare ciò, il gruppo acetil acetilico viene trasferito al gruppo TIOL (-sh) di uno dei residui di cisteina della subunità ACP di acidi grassi sintasi. Questa reazione è catalizzata dalla subunità ACP-Acil-transferasi.
Il gruppo acetilico viene quindi trasferito dall'ACP a un altro residuo di cisteina nel sito catalitico di un'altra subunità enzimatica del complesso che è noto come β-cethoacil-ACP-SINTA. Pertanto, il complesso enzimatico è "orzo" per iniziare la sintesi.
Passaggio 2: trasferimento di unità malonil-coa
Il malonil-CoA prodotto dal carbossilasa acetil-CoA viene trasferito al gruppo Tiol nell'ACP e durante questa reazione si perde la porzione di COA. La reazione è catalizzata dalla subunità malonil-ACP-transferasi del complesso di acidi grassi sintasi, che quindi produce malonil-ACP.
Durante questo processo, il gruppo malonico è collegato all'ACP e al β-cotoacil-ACP-sita attraverso un collegamento estere e un altro solfidl, rispettivamente.
Passaggio 3: condensa
L'enzima β-Zoacil-ACP-Suntase catalizza il trasferimento del gruppo acetilico che si è unito a lui nel passaggio di "orzo" verso il carbonio 2 del gruppo Malonilo che, nel passaggio precedente, è stato trasferito all'ACP.
Durante questa reazione viene rilasciata una molecola di CO2 che proviene dal malonio, che corrisponde alla CO2 fornita dal bicarbonato nella reazione carbossilazione della carbossilasi acetil-CoA. Quindi, si verifica acetoacetil-ACP.
Passaggio 4: riduzione
La subunità β-Zoacil-ACP-reduttasi catalizza la riduzione dell'aceoacetil-ACP dipendente da NADPH.
Passaggio 5: disidratazione
Questo passaggio forma il trans-α, β-acil-ACP o ∆2-Insaturado-acil-ACP (CRRATONIL-ACP), prodotto della disidratazione del d-β-idrossibutiril-ACP mediante azione dell'ACPRUNIT-ACPRUNIT -Hydratasa.
Più tardi, il cratonil-ACP è ridotto a butiril-ACP da una reazione dipendente dalla NADPH catalizzata dalla subunità arrabbiata-AC-reduttasi. Con questa reazione il primo dei sette cicli necessari per produrre palmitail-ACP, che è il precursore di quasi tutti gli acidi grassi è completato.
Come seguono le successive reazioni di condensa?
Il gruppo Butirilo viene trasferito dall'ACP al gruppo Tiol di un residuo di cisteina nel β-cethoacil-ACP-sita, in modo che l'ACP sia in grado di accettare un altro gruppo malonico dal malonil-CoA.
In questo modo, la reazione che accade è la condensa di Malonil-ACP con Buturil-β-Cotoacil-ACP-Sinta, che dà origine a β-zotohexanoil-ACP + CO2.
L'ACP di Palmitail che deriva dai successivi passaggi (dopo l'aggiunta di 5 unità più maloniche) può essere rilasciata come acido palmitico libero grazie all'attività dell'enzima tioesterasi, può essere trasferito al COA o incorporare nell'acido fosfatidico per La via di sintesi di fosfolipidi e triacilgliceridi.
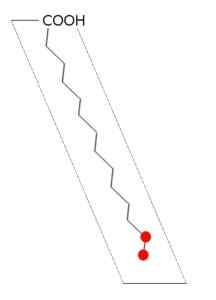
 Struttura dell'acido palmitico (fonte: Andel, via Wikimedia Commons)
Struttura dell'acido palmitico (fonte: Andel, via Wikimedia Commons) La sintasi degli acidi grassi della maggior parte degli organismi si ferma nella sintesi di Palmitail-ACP, dato che il sito catalitico della subunità β-cethoacil-ACP-sintasi ha una configurazione in cui solo gli acidi grassi di quella lunghezza possono essere alloggiati.
In che modo sono gli acidi grassi di un numero dispari di atomi di carbonio?
Questi sono relativamente comuni negli organismi marini e sono anche sintetizzati da un complesso acido grasso sintasi. Tuttavia, la reazione "orzo" si verifica con una molecola più lunga, il propionil-ACP, di tre atomi di carbonio.
Dove e come sono gli acidi grassi delle catene più lunghe?
L'acido palmitico, come commentato, funge da precursore per molti acidi grassi saturi e insaturi di catene più lunghe. Il processo di "allungamento" degli acidi grassi si verifica nei mitocondri, mentre l'introduzione dell'insaturazione si verifica essenzialmente nel reticolo endoplasmatico.
Molti organismi trasformano i loro acidi grassi saturi in insaturi come adattamento a basse temperature ambientali, poiché ciò consente loro di mantenere il punto di fusione dei lipidi al di sotto della temperatura ambiente.
Proprietà degli acidi grassi
Molte proprietà degli acidi grassi dipendono dalla loro lunghezza della catena e dalla presenza e dal numero di insaturazione:
Può servirti: teoria della sintesi abiotica: caratteristiche principali- Gli acidi grassi insaturi hanno punti di fusione più bassi rispetto agli acidi grassi saturi della stessa lunghezza.
- La lunghezza degli acidi grassi (il numero di atomi di carbonio) è inversamente proporzionale alla fluidità o alla flessibilità della molecola, cioè le molecole più "corte" sono più fluide e viceversa.
In generale, le sostanze grasse fluide sono composte da acidi grassi con catene corte e con insaturazione presenza.
Le piante hanno una quantità abbondante di acidi grassi insaturi, così come animali che vivono a temperature molto basse, poiché questi, come componenti dei lipidi presenti nelle membrane cellulari, danno loro una maggiore fluidità in queste condizioni.
In condizioni fisiologiche, la presenza di un doppio legame nella catena di idrocarburi di un grasso.
La presenza di doppi legami negli acidi grassi associati alle molecole lipidiche ha effetti diretti sul grado di "imballaggio" che possono avere nelle membrane a cui appartengono e con esso hanno anche effetti sulle proteine della membrana.
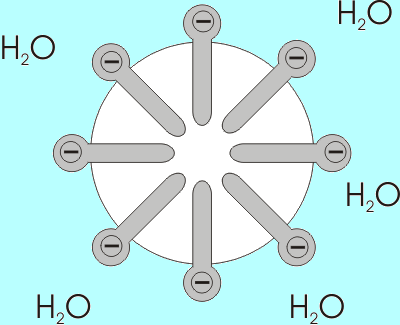 Esempio della formazione di un Myla di acidi grassi con i gruppi carbossilici esposti all'ambiente acquoso (Fonte: Benutzer: Anderl [CC BY-SA 3.0 (http: // creativeCommons.Org/licenze/by-sa/3.0/)] via Wikimedia Commons)
Esempio della formazione di un Myla di acidi grassi con i gruppi carbossilici esposti all'ambiente acquoso (Fonte: Benutzer: Anderl [CC BY-SA 3.0 (http: // creativeCommons.Org/licenze/by-sa/3.0/)] via Wikimedia Commons) La solubilità degli acidi grassi diminuisce all'aumentare della lunghezza della sua catena, quindi sono inversamente proporzionali. Nelle miscele acquose e lipidiche, gli acidi grassi sono associati nelle strutture note come micelle.
Una micella è una struttura in cui le catene alifatiche di acidi grassi sono "racchiuse", quindi "espellendo" tutte le molecole d'acqua e sulla cui superficie ci sono gruppi di carbossili.
Nomenclatura
La nomenclatura degli acidi grassi può essere in qualche modo complessa, soprattutto se si riferisce ai nomi comuni che ricevono, che sono spesso correlati ad alcune proprietà fisico -chimiche, con il luogo in cui si trovano o altre caratteristiche.
Molti autori considerano che grazie al gruppo carbossilico terminale queste molecole sono ionizzate a pH fisiologico, si dovrebbe riferirsi ad essi come "carbossilati" usando la terminazione "Ato ".
Secondo il sistema Iupac, l'enumerazione degli atomi di carbonio di un grasso. Il metil del terminale a catena contiene l'atomo di carbonio ω.
In generale, nella nomenclatura sistematica viene dato il nome dell'idrocarburo "parentale" (idrocarburo con lo stesso numero di atomi di carbonio) e la loro terminazione viene sostituita "O" di "Oico", Se si tratta di un acido grasso insaturo, viene aggiunta la terminazione "Nebbia".
Considera, ad esempio, il caso di un acido grasso di 18 atomi di carbonio (C18):
- Poiché l'idrocarburo con la stessa quantità di atomi di carbonio è noto come Octadecano, l'acido saturo è chiamato "Acido ottadecanoico"O bene"Octadecanoato"E il suo nome comune è l'acido stearico.
- Se hai un doppio legame tra una coppia di atomi di carbonio nella sua struttura, è noto come "Acido ottadecenoico"
- Se hai due doppie c - c - c - allora viene chiamato "Acido ottadecadienoico" E se ne hai tre "Acido ottadecatrienoico".
Se si desidera riassumere la nomenclatura, utilizzare 18: 0 per acido grasso a 18 carbonio e nessun doppio legame (saturo) e, a seconda del grado di insaturazione, quindi invece di zero è scritto 18: 1 per una molecola con una molecola con insaturazione, 18: 2 per uno con due insaturazioni e così via.
Se si desidera specificare tra quali atomi di carbonio sono i doppi legami negli acidi grassi insaturi, il simbolo ∆ viene utilizzato con un sondaggio numerico che indica il luogo di insaturazione e prefisso "Cis" O "trans", a seconda della configurazione di questo.
Riferimenti
- Badui, s. (2006). Chimica alimentare. (E. Quintanar, ed.) (4a ed.). Messico d.F.: Pearson Education.
- Garrett, r., & Grisham, C. (2010). Biochimica (4a ed.). Boston, USA: Brooks/Cole. Apprendimento del Cengage.
- Mathews, c., Van Holde, K., & Ahern, K. (2000). Biochimica (3 ° ed.). San Francisco, California: Pearson.
- Murray, r., Bender, d., Botham, k., Kennelly, p., Rodwell, v., & Weil, p. (2009). Biochimica illustrata di Harper (28 ° ed.). McGraw-Hill Medical.
- Nelson, d. L., & Cox, m. M. (2009). Principi di biochimica Lechinger. Omega Editions (5 ° ed.).
- Rawn, j. D. (1998). Biochimica. Burlington, Massachusetts: Neil Patterson Publishers.
- TVRZICKA, E., Kremmyda, l., Stankova, b., & Zak, a. (2011). Acidi grassi come biocompounds: il loro ruolo nel metabolismo umano, nella salute e nelle malattie: una revisione. Parte 1: classificazione, fonti dietetiche e funzioni biologiche. Biomed Pap Med Fac Univ Palacky Olomouc ceco Repub, 155(2), 117-130.
- « Struttura, funzioni, classi ed esempi di glicoproteine
- L'impatto dell'appartenenza alla cittadinanza digitale »