

Sfingolipidi Cosa sono, caratteristiche, funzioni, sintesi
- 779
- 89
- Zelida Gatti
Cosa sono gli sfilulipidi?
IL sfingolipidi Rappresentano una delle tre grandi famiglie lipidiche presenti nelle membrane biologiche. Come glyceoposfolipidi e steroli, sono molecole anfipatiche con una regione polare idrofila e una regione apolare idrofobica.
Furono descritti per la prima volta nel 1884 da Johann L. W. Thudichum, che ha descritto tre sfingolipidi (sfingomyeline, cerebrale e cervello -cerve.
A differenza dei glyceofosfolipidi, gli sfingolipidi non sono costruiti su una molecola di glicerolo a 3 fosfato come scheletro principale, ma sono composti derivati da Sphiny, un aminoalcool con una lunga catena di idrocarburi attaccata da un legame Amida.
EStruttura degli sfingolipidi
Tutti gli sfingolipidi derivano da una serina L, che è condensato con un acido grasso a catena lunga per formare la base sfingoidica, nota anche come base a catena lunga (LCB).
Le basi più comuni sono la sfinganina e la sfinxina, che differiscono l'una dall'altra in presenza di un doppio legame trans Tra i carboni 4 e 5 dell'acido grasso della sfinxina.
I carboni 1, 2 e 3 della sfinxina sono strutturalmente analoghi ai glicerofosfolipidi glicerolo glicerolo. Quando i legami di Amida lega un acido grasso al carbonio 2 della gravidanza.
Gli acidi grassi a catena lunga che costituiscono le regioni idrofobiche di questi lipidi possono essere molto diversi. Le lunghezze variano da 14 a 22 atomi di carbonio che possono avere diversi gradi di saturazione, di solito tra i carboni 4 e 5.
Nelle posizioni 4 o 6 possono avere gruppi di idrossilli e doppi legami in altre posizioni o persino ramificazioni come i gruppi metilici.
Caratteristiche
Le catene di acidi grassi collegati dai legami di Amida alle ceramidi sono comunemente saturi e tendono ad avere una lunghezza maggiore di quelle trovate nei glicerofosfolipidi, che sembrano essere cruciali per l'attività biologica di questi.
Una caratteristica distintiva dello scheletro sfingolipide è che possono avere un carico netto positivo a pH neutro, strano tra le molecole lipidiche.
Comunque, il PkA del gruppo amminico è basso rispetto a una semplice ammina, tra 7 e 8, quindi una parte della molecola non viene caricata al pH fisiologico, il che potrebbe spiegare il movimento "libero" di questi tra i bilapas.
La tradizionale classificazione degli sfingolipidi deriva dalle molteplici modifiche che la molecola di ceramide può soffrire, specialmente in termini di sostituzioni dei gruppi di testa polare.
Può servirti: epadnavirus: caratteristiche, morfologia, trattamentoFunzioni
Gli sfingolipidi sono essenziali in animali, piante e funghi, nonché in alcuni organismi e virus procariotici.
-Funzioni strutturali
Gli sfingolipidi modulano le proprietà fisiche delle membrane, compresa la loro fluidità, spessore e curvatura. Modulari queste proprietà danno anche loro un'influenza diretta sull'organizzazione spaziale delle proteine della membrana.
In "zattere" lipidiche
Nelle membrane biologiche, i micro domini dinamici possono essere rilevati con una fluidità più bassa formata da molecole di colesterolo e sfingolipidi chiamate balsas lipidiche.
Queste strutture si verificano naturalmente e mantengono una stretta relazione con proteine complete, recettori della superficie cellulare e proteine di segnalazione, trasportatori e altre proteine con ancore di glicosilfosfatidilinitolo (GPI) (GPI).
-Funzioni di segnalazione
Hanno funzioni come molecole di segnalazione che fungono da secondi messaggeri o come ligandi secreti per i recettori della superficie cellulare.
Come messaggeri secondari possono partecipare alla regolazione dell'omeostasi del calcio, della crescita cellulare, della tumorogenesi e della soppressione dell'apoptosi. Inoltre, l'attività di molte proteine di membrana integrale e periferica dipende dalla sua associazione con gli sfingolipidi.
Molte interazioni intercellulari e cellulari con l'ambiente circostante dipendono dall'esposizione dei diversi gruppi polari degli sfingolipidi verso la faccia esterna della membrana plasmatica.
L'unione di glicosfingolipidi e lectina è cruciale per l'associazione della mielina con gli assoni, l'adesione dei neutrofili all'endotelio, ecc.
Dai prodotti del suo metabolismo
Gli sfingolipidi di segnalazione più importanti sono le basi a catena lunga o le sfingi e le ceramidi, nonché i loro derivati fosforilati, come la sfidosina 1-fosfato.
I prodotti del metabolismo di molti sfingolipidi attivano o inibiscono multipli più bianchi (proteine della chinasi, fosfatosio e altri), che controllano comportamenti cellulari complessi come crescita, differenziazione e apoptosi.
-Come recettori della membrana
Alcuni agenti patogeni usano Gloysphingolipidi come recettori per mediare il loro ingresso alle cellule ospiti o per offrire fattori di virulenza ad essi.
È stato dimostrato che gli sfidollipidi partecipano a più eventi cellulari come secrezione, endocitosi, chimico, neurotrasmissione, angiogenesi e infiammazione.
Sono anche coinvolti nel traffico della membrana, quindi influenzano l'internalizzazione di recettori, ordine, movimento e fusione di vescicole secretorie in risposta a diversi stimoli.
Gruppi sfingolipidi
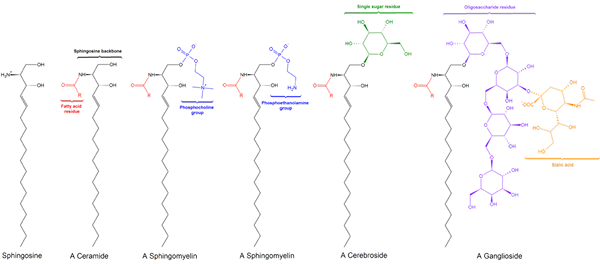
Esistono tre sottoclassi sfingolipidi, tutti derivati dalla ceramide e che differiscono l'uno dall'altro dai gruppi polari, vale a dire: sfilulomilins, glicolipidi e gangliari.
Sfingomielinas
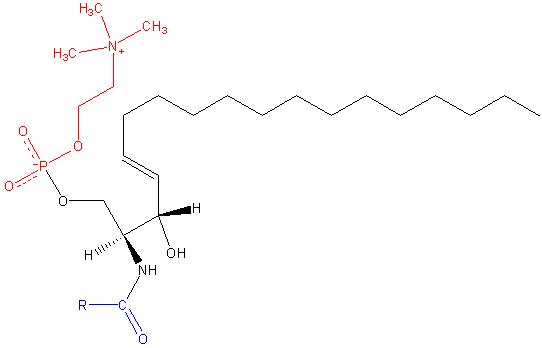 Sfingomileina. Nero: sfinxina. Rosso: fosfocolina. Blu: acido grasso.
Sfingomileina. Nero: sfinxina. Rosso: fosfocolina. Blu: acido grasso. Questi contengono fosfocolina o fosfoetanolamina come gruppo di testa polare, quindi sono classificati come fosfolipidi insieme a glyceoposfolipidi. Sembrano, ovviamente, fosfatidilcoline in struttura a tre dimensioni e proprietà generali poiché non hanno alcuna carica nelle loro teste polari.
Può servirti: enolasi: struttura, meccanismo d'azione, funzioniSono presenti nelle membrane plasmatiche delle cellule animali e sono particolarmente abbondanti nella mielina, un baccello che circonda e isola gli assoni di alcuni neuroni.
Glicolipidi o glicosfingolipidi neutri (senza carico)
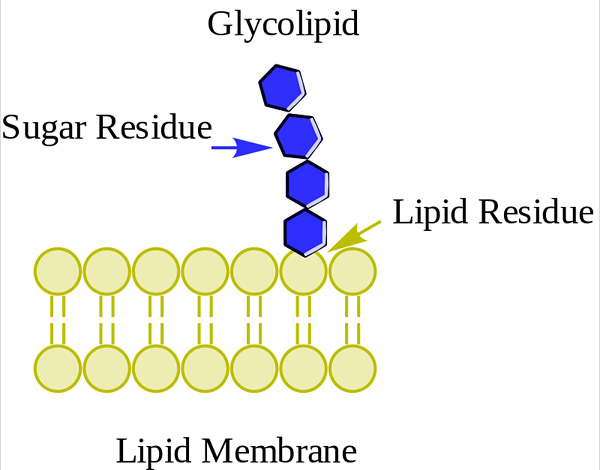 Glucolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)], da Wikimedia Commons
Glucolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licenze/by-sa/4.0)], da Wikimedia Commons Sono principalmente sulla faccia esterna della membrana plasmatica e hanno uno o più zuccheri come gruppo di testa polare direttamente uniti alla porzione di ceramide di carbonio 1 della porzione di ceramide. Non hanno gruppi di fosfato. Poiché il ph 7 non ha carico, sono chiamati glicolipidi neutri.
I cerebrosidi hanno una singola molecola di zucchero collegata alla ceramide. Quelli contenenti galattosio si trovano nelle membrane plasmatiche delle cellule tissutali non nerve. I globosidi sono goustoesphingolipidi con due o più zuccheri, normalmente D-glicosio, D-galattosio o N-acetil-D-galattosamina.
Gangliósidos o glicosfingolipidi
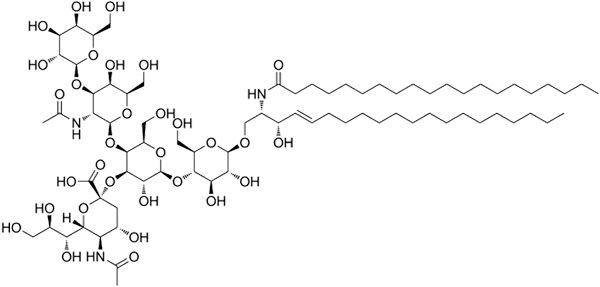 Struttura gangloside GM1
Struttura gangloside GM1 Questi sono gli sfingolipidi più complessi. Hanno oligosaccaridi come gruppo di testa polare e uno o più residui di acido n-acetilmuramico terminale, chiamati anche acido siral. L'acido siamico porta ai gangli il carico negativo a pH 7, che li distingue dai glicosfingolipidi neutri.
La nomenclatura di questo tipo di sfingolipidi dipende dalla quantità di rifiuti di acido sinalico presenti nella parte oligosaccaridica della testa polare.
Sintesi
La molecola di base a catena lunga o sfinina viene sintetizzata nel reticolo endoplasmatico (ER) e l'aggiunta del gruppo polare a testa di questi lipidi si verifica successivamente nel complesso del Golgi. Nei mammiferi, nei mitocondri può verificarsi anche una sintesi di sfingolipidi.
Dopo aver completato la loro sintesi nel complesso del Golgi, gli sfingolipidi vengono trasportati in altri scomparti cellulari attraverso meccanismi mediati dalle vescicole.
La biosintesi degli sfingolipidi è costituita da tre eventi fondamentali: la sintesi delle basi a catena lunga, la biosintesi delle ceramidi da parte dell'Unione di un acido grasso attraverso un legame Amida e, infine, la formazione degli sfingolipidi complessi con mezzi dell'Unione dell'Unione di gruppi polari nel carbonio 1 della base di sfingoide.
Oltre alla sintesi di novo, Gli sfingolipidi possono anche formarsi per sostituzione o riciclaggio delle basi e delle ceramidi a catena lunga, che possono alimentare il pool di sfingolipidi.
Sintesi di scheletro in ceramide
La biosintesi della ceramide, lo scheletro degli sfingolipidi, inizia con la condensazione decarbossilerativa di una molecola di palmotail-CoA e una l-serina. La reazione è catalizzata da una serina palmotail transferasi (SPT) eterodimerica, fosfato piridossale e il prodotto è il 3-zo diidroesfingosina ze.
Può servirti: microbiota: composizione, funzioni e dove si trovaQuesto enzima è inibito da β-faal-L-alanine e l-cicloserinas. Nei lieviti è codificato da due geni, mentre nei mammiferi ci sono tre geni per questo enzima. Il sito attivo si trova nella faccia citoplasmatica del reticolo endoplasmatico.
Il ruolo di questo primo enzima è preservato in tutti gli organismi studiati. Tuttavia, ci sono alcune differenze tra i taxa che hanno a che fare con la posizione subcellulare dell'enzima: i batteri sono citoplasmatici, lieviti, piante e animali sono nel reticolo endoplasmatico.
La 3-Zoesfinganina viene successivamente ridotta dalla 3-ketosfinganina dipendente da NADPH. Diidroceramide sintasi (sfinganina n-acil transferasi) acetila quindi sphinanina per produrre diidroceramide. La ceramide viene quindi formata dalla diidroceramide deatturasi/reduttasi, che inserisce un doppio legame trans Nella posizione 4-5.
Nei mammiferi ci sono numerose sintasi di isoforme in ceramide, ognuna delle quali si unisce a una specifica catena di acidi grassi alle basi delle catene lunghe. Pertanto, le ceramidi sintesi e altri enzimi, Elongasas, forniscono la principale fonte di diversità di acidi grassi negli sfingolipidi.
Metabolismo
La degradazione degli sfingolipidi viene eseguita dagli enzimi glihochydico e sfingomielinasi, che sono responsabili della rimozione delle modifiche dei gruppi polari. D'altra parte, le ceramidasi rigenerano le basi a catena lunghe dalle ceramidi.
I gangli sono degradati da un insieme di enzimi lisosomiali che catalizzano il passo dopo passo delle unità di zucchero, producendo una ceramide alla fine.
Un altro modo di degradazione è l'internalizzazione degli sfingolipidi nelle vescicole endocitiche che vengono inviate alla membrana plasmatica o trasportate su lisosomi dove sono degradati da specifica idrolasi acida.
Non tutte le basi a catena lunghe vengono riciclate, il mantenimento endoplasmatico. Questo meccanismo di degradazione consiste nella fosforilazione anziché dalla acilazione degli LCB, dando origine a molecole di segnalazione che possono essere substrati solubili per l'enzima liases.
Regolamento
Il metabolismo di questi lipidi è regolato su diversi livelli, uno di questi è quello degli enzimi responsabili della sintesi, le loro modifiche post -traduzionali e gli stessi meccanismi degli stessi.
Alcuni meccanismi di regolazione sono specifiche cellulari, sia per controllare il tempo di sviluppo cellulare in cui vengono prodotti o in risposta a segnali specifici.
Riferimenti
- Bartke, n., & Hannun, e. (2009). SPPHOLIPIDI Bioattivi: metabolismo e funzione. Journal of Lipid Research, cinquanta, 19.
- Breslow, d. K. (2013). Omeostasi sfingolipidica nel reticolo endoplasmatico e oltre. Prospettive di Cold Spring Harbor in biologia, 5(4), A013326.